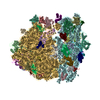
ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 3jbu | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | Mechanisms of Ribosome Stalling by SecM at Multiple Elongation Steps | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | RIBOSOME / single particle analysis / ribosome stalling | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報outer membrane protein complex / monoatomic ion transmembrane transporter activity / detection of virus / outer membrane / porin activity / pore complex / ornithine decarboxylase inhibitor activity / transcription antitermination factor activity, RNA binding / misfolded RNA binding / Group I intron splicing ...outer membrane protein complex / monoatomic ion transmembrane transporter activity / detection of virus / outer membrane / porin activity / pore complex / ornithine decarboxylase inhibitor activity / transcription antitermination factor activity, RNA binding / misfolded RNA binding / Group I intron splicing / RNA folding / transcriptional attenuation / endoribonuclease inhibitor activity / RNA-binding transcription regulator activity / positive regulation of ribosome biogenesis / negative regulation of cytoplasmic translation / four-way junction DNA binding / monoatomic ion transport / DnaA-L2 complex / translation repressor activity / negative regulation of translational initiation / regulation of mRNA stability / negative regulation of DNA-templated DNA replication initiation / mRNA regulatory element binding translation repressor activity / assembly of large subunit precursor of preribosome / positive regulation of RNA splicing / ribosome assembly / transcription elongation factor complex / regulation of DNA-templated transcription elongation / cytosolic ribosome assembly / response to reactive oxygen species / DNA endonuclease activity / transcription antitermination / regulation of cell growth / DNA-templated transcription termination / response to radiation / maintenance of translational fidelity / mRNA 5'-UTR binding / cell outer membrane / ribosome biogenesis / large ribosomal subunit / regulation of translation / ribosome binding / transferase activity / outer membrane-bounded periplasmic space / ribosomal small subunit biogenesis / ribosomal small subunit assembly / small ribosomal subunit / 5S rRNA binding / small ribosomal subunit rRNA binding / ribosomal large subunit assembly / cytosolic small ribosomal subunit / large ribosomal subunit rRNA binding / cytosolic large ribosomal subunit / cytoplasmic translation / tRNA binding / negative regulation of translation / rRNA binding / structural constituent of ribosome / ribosome / translation / response to antibiotic / negative regulation of DNA-templated transcription / mRNA binding / DNA damage response / symbiont entry into host cell / DNA binding / RNA binding / zinc ion binding / identical protein binding / membrane / cytosol / cytoplasm 類似検索 - 分子機能 | ||||||
| 生物種 |  | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.64 Å | ||||||
 データ登録者 データ登録者 | Zhang, J. / Pan, X.J. / Yan, K.G. / Sun, S. / Gao, N. / Sui, S.F. | ||||||
 引用 引用 | ジャーナル: Elife / 年: 2015 タイトル: Mechanisms of ribosome stalling by SecM at multiple elongation steps. 著者: Jun Zhang / Xijiang Pan / Kaige Yan / Shan Sun / Ning Gao / Sen-Fang Sui /  要旨: Regulation of translating ribosomes is a major component of gene expression control network. In Escherichia coli, ribosome stalling by the C-terminal arrest sequence of SecM regulates the SecA- ...Regulation of translating ribosomes is a major component of gene expression control network. In Escherichia coli, ribosome stalling by the C-terminal arrest sequence of SecM regulates the SecA-dependent secretion pathway. Previous studies reported many residues of SecM peptide and ribosome exit tunnel are critical for stalling. However, the underlying molecular mechanism is still not clear at the atomic level. Here, we present two cryo-EM structures of the SecM-stalled ribosomes at 3.3-3.7 Å resolution, which reveal two different stalling mechanisms at distinct elongation steps of the translation cycle: one is due to the inactivation of ribosomal peptidyl-transferase center which inhibits peptide bond formation with the incoming prolyl-tRNA; the other is the prolonged residence of the peptidyl-RNA at the hybrid A/P site which inhibits the full-scale tRNA translocation. These results demonstrate an elegant control of translation cycle by regulatory peptides through a continuous, dynamic reshaping of the functional center of the ribosome. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 3jbu.cif.gz | 3.5 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb3jbu.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 3jbu.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 3jbu_validation.pdf.gz | 1.5 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 3jbu_full_validation.pdf.gz | 1.7 MB | 表示 | |
| XML形式データ | 3jbu_validation.xml.gz | 202.3 KB | 表示 | |
| CIF形式データ | 3jbu_validation.cif.gz | 355.5 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/jb/3jbuftp://data.pdbj.org/pub/pdb/validation_reports/jb/3jbu | HTTPS FTP |
-関連構造データ







-リンク
 PDBj
PDBj






























- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-30S ribosomal protein ... , 20種, 20分子 BCDEFGHIJKLMNOPQRSTU
| #1: タンパク質 | 分子量: 26781.670 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #2: タンパク質 | 分子量: 26031.316 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #3: タンパク質 | 分子量: 23514.199 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #4: タンパク質 | 分子量: 17629.398 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #5: タンパク質 | 分子量: 15197.032 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #6: タンパク質 | 分子量: 17637.445 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #7: タンパク質 | 分子量: 14146.557 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #8: タンパク質 | 分子量: 14886.270 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #9: タンパク質 | 分子量: 11755.597 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #10: タンパク質 | 分子量: 13870.975 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #11: タンパク質 | 分子量: 13768.157 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #12: タンパク質 | 分子量: 13128.467 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #13: タンパク質 | 分子量: 11606.560 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #14: タンパク質 | 分子量: 10319.882 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #15: タンパク質 | 分子量: 9207.572 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #16: タンパク質 | 分子量: 9724.491 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #17: タンパク質 | 分子量: 9005.472 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #18: タンパク質 | 分子量: 10455.355 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #19: タンパク質 | 分子量: 9708.464 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #20: タンパク質 | 分子量: 8524.039 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
+50S ribosomal protein ... , 28種, 28分子 01234678cdefghjklmnopqrstuwy
-タンパク質 , 1種, 1分子 z
| #49: タンパク質 | 分子量: 9224.300 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|
-RNA鎖 , 5種, 5分子 AXabv
| #50: RNA鎖 | 分子量: 499690.031 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #51: RNA鎖 | 分子量: 3458.106 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #52: RNA鎖 | 分子量: 38790.090 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #53: RNA鎖 | 分子量: 941612.375 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #54: RNA鎖 | 分子量: 24490.562 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-詳細
| 配列の詳細 | THE SEQUENCE OF THE CHAIN Z (ENTITY 56, UNCAPITALIZED Z) WAS NOT AVAILABLE AT THE UNIPROT ...THE SEQUENCE OF THE CHAIN Z (ENTITY 56, UNCAPITALI |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: SecM-Gly-RNC / タイプ: RIBOSOME |
|---|---|
| 緩衝液 | 名称: 20mM HEPES, 50mM KOAc, 6mM Mg(OAc)2, 1mM DTT, 500 ug/ml chloramphenicol, 0.05% Nikkol, 0.5% pill/ml Complete EDTA-free Protease inhibitor cocktail, 0.1U/ml RNasin and 125mM sucrose pH: 7 詳細: 20mM HEPES, 50mM KOAc, 6mM Mg(OAc)2, 1mM DTT, 500 ug/ml chloramphenicol, 0.05% Nikkol, 0.5% pill/ml Complete EDTA-free Protease inhibitor cocktail, 0.1U/ml RNasin and 125mM sucrose |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 試料支持 | 詳細: Sample are made by Cryo-EM on the holy carbon |
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS / 日付: 2014年5月8日 |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 22500 X / 倍率(補正後): 37878 X / 最大 デフォーカス(公称値): 1000 nm / 最小 デフォーカス(公称値): 3500 nm / Cs: 2.7 mm |
| 撮影 | 電子線照射量: 16 e/Å2 フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) |
- 解析
解析
| EMソフトウェア |
| |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | 詳細: CTFFIND | |||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | |||||||||||||||
| 3次元再構成 | 手法: REFERENCE-BASED RECONSTRUCTION / 解像度: 3.64 Å / 粒子像の数: 60354 / ピクセルサイズ(公称値): 1.32 Å / ピクセルサイズ(実測値): 1.31 Å / 対称性のタイプ: POINT | |||||||||||||||
| 原子モデル構築 | プロトコル: FLEXIBLE FIT / 空間: RECIPROCAL / 詳細: METHOD--FLEXIBLE | |||||||||||||||
| 原子モデル構築 | PDB-ID: 4V7T Accession code: 4V7T / Source name: PDB / タイプ: experimental model | |||||||||||||||
| 精密化 | 最高解像度: 3.64 Å | |||||||||||||||
| 精密化ステップ | サイクル: LAST / 最高解像度: 3.64 Å
|