Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-3j8y: High-resolution structure of ATP analog-bound kinesin on microtubules -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 3j8y | ||||||
|---|---|---|---|---|---|---|---|
| Title | High-resolution structure of ATP analog-bound kinesin on microtubules | ||||||
 Components Components |
| ||||||
 Keywords Keywords | MOTOR PROTEIN/STRUCTURAL PROTEIN / molecular motors / kinesin / myosin / microtubules / cytoskeletal motors / MOTOR PROTEIN-STRUCTURAL PROTEIN complex | ||||||
| Function / homology |  Function and homology information Function and homology informationregulation of modification of synapse structure, modulating synaptic transmission / plus-end-directed vesicle transport along microtubule / cytolytic granule membrane / cytoplasm organization / anterograde dendritic transport of neurotransmitter receptor complex / mitocytosis / anterograde axonal protein transport / anterograde neuronal dense core vesicle transport / retrograde neuronal dense core vesicle transport / ciliary rootlet ...regulation of modification of synapse structure, modulating synaptic transmission / plus-end-directed vesicle transport along microtubule / cytolytic granule membrane / cytoplasm organization / anterograde dendritic transport of neurotransmitter receptor complex / mitocytosis / anterograde axonal protein transport / anterograde neuronal dense core vesicle transport / retrograde neuronal dense core vesicle transport / ciliary rootlet / lysosome localization / positive regulation of potassium ion transport / plus-end-directed microtubule motor activity / RHO GTPases activate KTN1 / Kinesins / vesicle transport along microtubule / Microtubule-dependent trafficking of connexons from Golgi to the plasma membrane / Resolution of Sister Chromatid Cohesion / Hedgehog 'off' state / Cilium Assembly / Intraflagellar transport / COPI-dependent Golgi-to-ER retrograde traffic / Mitotic Prometaphase / Carboxyterminal post-translational modifications of tubulin / RHOH GTPase cycle / EML4 and NUDC in mitotic spindle formation / Sealing of the nuclear envelope (NE) by ESCRT-III / Kinesins / PKR-mediated signaling / Separation of Sister Chromatids / The role of GTSE1 in G2/M progression after G2 checkpoint / Aggrephagy / RHO GTPases activate IQGAPs / RHO GTPases Activate Formins / HSP90 chaperone cycle for steroid hormone receptors (SHR) in the presence of ligand / centrosome localization / MHC class II antigen presentation / Recruitment of NuMA to mitotic centrosomes / microtubule motor activity / kinesin complex / COPI-mediated anterograde transport / mitochondrion transport along microtubule / COPI-dependent Golgi-to-ER retrograde traffic / microtubule-based movement / natural killer cell mediated cytotoxicity / stress granule disassembly / Insulin processing / synaptic vesicle transport / postsynaptic cytosol / microtubule-based process / phagocytic vesicle / axon cytoplasm / MHC class II antigen presentation / dendrite cytoplasm / axon guidance / positive regulation of synaptic transmission, GABAergic / positive regulation of protein localization to plasma membrane / sperm end piece / regulation of membrane potential / structural constituent of cytoskeleton / cellular response to type II interferon / microtubule cytoskeleton organization / centriolar satellite / Signaling by ALK fusions and activated point mutants / mitotic cell cycle / microtubule cytoskeleton / nuclear membrane / microtubule binding / vesicle / microtubule / Hydrolases; Acting on acid anhydrides; Acting on GTP to facilitate cellular and subcellular movement / cadherin binding / GTPase activity / GTP binding / protein-containing complex binding / perinuclear region of cytoplasm / ATP hydrolysis activity / mitochondrion / ATP binding / membrane / metal ion binding / identical protein binding / cytoplasm / cytosol Similarity search - Function | ||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||
| Method | ELECTRON MICROSCOPY / helical reconstruction / cryo EM / Resolution: 5 Å | ||||||
 Authors Authors | Shang, Z. / Zhou, K. / Xu, C. / Csencsits, R. / Cochran, J.C. / Sindelar, C.V. | ||||||
 Citation Citation | Journal: Elife / Year: 2014 Title: High-resolution structures of kinesin on microtubules provide a basis for nucleotide-gated force-generation. Authors: Zhiguo Shang / Kaifeng Zhou / Chen Xu / Roseann Csencsits / Jared C Cochran / Charles V Sindelar /  Abstract: Microtubule-based transport by the kinesin motors, powered by ATP hydrolysis, is essential for a wide range of vital processes in eukaryotes. We obtained insight into this process by developing ...Microtubule-based transport by the kinesin motors, powered by ATP hydrolysis, is essential for a wide range of vital processes in eukaryotes. We obtained insight into this process by developing atomic models for no-nucleotide and ATP states of the monomeric kinesin motor domain on microtubules from cryo-EM reconstructions at 5-6 Å resolution. By comparing these models with existing X-ray structures of ADP-bound kinesin, we infer a mechanistic scheme in which microtubule attachment, mediated by a universally conserved 'linchpin' residue in kinesin (N255), triggers a clamshell opening of the nucleotide cleft and accompanying release of ADP. Binding of ATP re-closes the cleft in a manner that tightly couples to translocation of cargo, via kinesin's 'neck linker' element. These structural transitions are reminiscent of the analogous nucleotide-exchange steps in the myosin and F1-ATPase motors and inform how the two heads of a kinesin dimer 'gate' each other to promote coordinated stepping along microtubules. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 3j8y.cif.gz | 225.9 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb3j8y.ent.gz | 169.2 KB | Display | PDB format |
| PDBx/mmJSON format | 3j8y.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/j8/3j8yftp://data.pdbj.org/pub/pdb/validation_reports/j8/3j8y | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  6188MC  6187C  3j8xC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |







-Links
 PDBj
PDBj











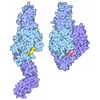


- Assembly
Assembly
| Deposited unit | 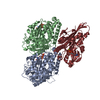
|
|---|---|
| 1 |
|
| Symmetry | Helical symmetry: (Circular symmetry: 1 / Dyad axis: no / N subunits divisor: 1 / Num. of operations: 1 / Rise per n subunits: 8.5215 Å / Rotation per n subunits: -25.77 °) |
| Details | The reconstructed 14-protofilament microtubule is pseudo-symmetric, containing a seam with 3 starts per tubulin monomer, or 1.5 starts per tubulin dimer. |
-Components
-Protein , 3 types, 3 molecules KAB
| #1: Protein | Mass: 39238.145 Da / Num. of mol.: 1 Fragment: Truncated catalytic head domain (monomeric, UNP residues 1-349) Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: KIF5B, KNS, KNS1 / Production host:  |
|---|---|
| #2: Protein | Mass: 50204.445 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #3: Protein | Mass: 49983.824 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
-Non-polymers , 4 types, 4 molecules 

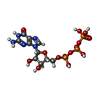
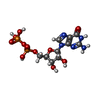
| #4: Chemical | ChemComp-ATP /  Mass: 507.181 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Comment: ATP, energy-carrying molecule*YM Mass: 507.181 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Comment: ATP, energy-carrying molecule*YM |
|---|---|
| #5: Chemical | ChemComp-MG /  Mass: 24.305 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Mg |
| #6: Chemical | ChemComp-GTP /  Mass: 523.180 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H16N5O14P3 / Comment: GTP, energy-carrying molecule*YM Mass: 523.180 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H16N5O14P3 / Comment: GTP, energy-carrying molecule*YM |
| #7: Chemical | ChemComp-GDP /  Type: RNA linking / Mass: 443.201 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H15N5O11P2 / Comment: GDP, energy-carrying molecule*YM Type: RNA linking / Mass: 443.201 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H15N5O11P2 / Comment: GDP, energy-carrying molecule*YM |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: FILAMENT / 3D reconstruction method: helical reconstruction |
- Sample preparation
Sample preparation
| Component |
| |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Value: 0.135 MDa / Experimental value: NO | |||||||||||||||||||||||||
| Buffer solution | Name: 25 mM PIPES, 25 mM NaCl, 2 mM MgCl2, 1 mM EGTA / pH: 6.8 / Details: 25 mM PIPES, 25 mM NaCl, 2 mM MgCl2, 1 mM EGTA | |||||||||||||||||||||||||
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | |||||||||||||||||||||||||
| Specimen support | Details: 300 mesh copper grid with homemade holey carbon | |||||||||||||||||||||||||
| Vitrification | Instrument: HOMEMADE PLUNGER / Cryogen name: ETHANE Details: No glow discharge was applied. After sample application to grid, liquid was mostly 'wicked' away by edgewise application of filter paper. Subsequently, blotting and plunge freezing were ...Details: No glow discharge was applied. After sample application to grid, liquid was mostly 'wicked' away by edgewise application of filter paper. Subsequently, blotting and plunge freezing were performed with ~0.5 second delay after blotting but prior to plunging into liquid ethane. Method: No glow discharge was applied. After sample application to grid, liquid was mostly 'wicked' away by edgewise application of filter paper. Subsequently, blotting and plunge freezing were ...Method: No glow discharge was applied. After sample application to grid, liquid was mostly 'wicked' away by edgewise application of filter paper. Subsequently, blotting and plunge freezing were performed with ~0.5 second delay after blotting but prior to plunging. |
- Electron microscopy imaging
Electron microscopy imaging
| Microscopy | Model: FEI TITAN / Date: Jun 2, 2013 Details: 4K x 4K counting mode was used. 24 frames total were collected. |
|---|---|
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Calibrated magnification: 23859 X / Nominal defocus max: 2500 nm / Nominal defocus min: 1000 nm / Cs: 2 mm / Camera length: 0 mm |
| Specimen holder | Specimen holder model: GATAN LIQUID NITROGEN |
| Image recording | Electron dose: 15 e/Å2 / Film or detector model: GATAN ULTRASCAN 4000 (4k x 4k) |
| Image scans | Num. digital images: 51 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Relative weight: 1 |
- Processing
Processing
| EM software |
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Details: done within FREALIGN | ||||||||||||
| Helical symmerty | Angular rotation/subunit: 25.77 ° / Axial rise/subunit: 8.5215 Å / Axial symmetry: C1 Details: The reconstructed 14-protofilament microtubule is pseudo-symmetric, containing a seam with 3 starts per tubulin monomer, or 1.5 starts per tubulin dimer. | ||||||||||||
| 3D reconstruction | Method: Single particle / Resolution: 5 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 49961 / Nominal pixel size: 2.097 Å / Actual pixel size: 2.097 Å Details: Initial alignment was done using customized SPIDER scripts. Reconstruction and subsequent refinement were done by FREALIGN. Symmetry type: HELICAL | ||||||||||||
| Atomic model building | Protocol: FLEXIBLE FIT / Space: REAL Target criteria: RMSD from the starting structure was monitored for convergence. Details: REFINEMENT PROTOCOL--flexible DETAILS--MDFF was performed using explicit solvation, after placing active-site water coordinates identified in high-resolution crystal structures of kinesins ...Details: REFINEMENT PROTOCOL--flexible DETAILS--MDFF was performed using explicit solvation, after placing active-site water coordinates identified in high-resolution crystal structures of kinesins ATP-like state. Side chains were excluded from the MDFF target potential. Following several equilibration steps, the relative strength of the EM map potential (GSCALE term) was slowly increased from 0 to 1 over the course of 10 nanoseconds. The t = 1.4 ns time point was selected to represent the final fitted model, based on the approximate convergence of the RMSD from the starting structure. | ||||||||||||
| Atomic model building | 3D fitting-ID: 1 / Accession code: 4HNA / Initial refinement model-ID: 1 / PDB-ID: 4HNA / Source name: PDB / Type: experimental model
| ||||||||||||
| Refinement step | Cycle: LAST
|