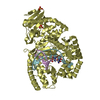
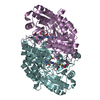
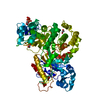
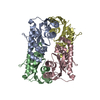
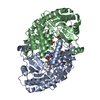
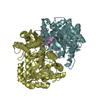
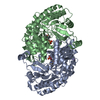
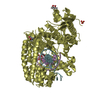
Journal: EMBO J / Year: 2008 Title: Molecular structure of the ParM polymer and the mechanism leading to its nucleotide-driven dynamic instability. Authors: David Popp / Akihiro Narita / Toshiro Oda / Tetsuro Fujisawa / Hiroshi Matsuo / Yasushi Nitanai / Mitsusada Iwasa / Kayo Maeda / Hirofumi Onishi / Yuichiro Maéda / Abstract: ParM is a prokaryotic actin homologue, which ensures even plasmid segregation before bacterial cell division. In vivo, ParM forms a labile filament bundle that is reminiscent of the more complex ...ParM is a prokaryotic actin homologue, which ensures even plasmid segregation before bacterial cell division. In vivo, ParM forms a labile filament bundle that is reminiscent of the more complex spindle formed by microtubules partitioning chromosomes in eukaryotic cells. However, little is known about the underlying structural mechanism of DNA segregation by ParM filaments and the accompanying dynamic instability. Our biochemical, TIRF microscopy and high-pressure SAX observations indicate that polymerization and disintegration of ParM filaments is driven by GTP rather than ATP and that ParM acts as a GTP-driven molecular switch similar to a G protein. Image analysis of electron micrographs reveals that the ParM filament is a left-handed helix, opposed to the right-handed actin polymer. Nevertheless, the intersubunit contacts are similar to those of actin. Our atomic model of the ParM-GMPPNP filament, which also fits well to X-ray fibre diffraction patterns from oriented gels, can explain why after nucleotide release, large conformational changes of the protomer lead to a breakage of intra- and interstrand interactions, and thus to the observed disintegration of the ParM filament after DNA segregation.
THE MOLECULE HAS MONOMERIC STATE AND FILAMENTOUS OLIGOMELIC STATE. PRECIOUS COORDINATES FOR A COMPLETE MULTIMER REPRESENTING THE KNOWN BIOLOGICALLY SIGNIFICANT OLIGOMERIZATION STATE OF THE MOLECULE HAS NOT YET BEEN KNOWN.
-
Components
#1: Protein
PlasmidsegregationproteinparM / Protein stbA / ParA locus 36 kDa protein
Mass: 35804.375 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Escherichia coli (E. coli) / Plasmid: PMD137 / Species (production host): Escherichia coli / Production host: Escherichia coli BL21 (bacteria) / Strain (production host): BL21 / References: UniProt: P11904
In the structure databanks used in Yorodumi, some data are registered as the other names, "COVID-19 virus" and "2019-nCoV". Here are the details of the virus and the list of structure data.
Jan 31, 2019. EMDB accession codes are about to change! (news from PDBe EMDB page)
EMDB accession codes are about to change! (news from PDBe EMDB page)
The allocation of 4 digits for EMDB accession codes will soon come to an end. Whilst these codes will remain in use, new EMDB accession codes will include an additional digit and will expand incrementally as the available range of codes is exhausted. The current 4-digit format prefixed with “EMD-” (i.e. EMD-XXXX) will advance to a 5-digit format (i.e. EMD-XXXXX), and so on. It is currently estimated that the 4-digit codes will be depleted around Spring 2019, at which point the 5-digit format will come into force.
The EM Navigator/Yorodumi systems omit the EMD- prefix.
Related info.:Q: What is EMD? / ID/Accession-code notation in Yorodumi/EM Navigator
Yorodumi is a browser for structure data from EMDB, PDB, SASBDB, etc.
This page is also the successor to EM Navigator detail page, and also detail information page/front-end page for Omokage search.
The word "yorodu" (or yorozu) is an old Japanese word meaning "ten thousand". "mi" (miru) is to see.
Related info.:EMDB / PDB / SASBDB / Comparison of 3 databanks / Yorodumi Search / Aug 31, 2016. New EM Navigator & Yorodumi / Yorodumi Papers / Jmol/JSmol / Function and homology information / Changes in new EM Navigator and Yorodumi
 Movie
Movie Controller
Controller


 Open data
Open data
 Basic information
Basic information Components
Components Keywords
Keywords Function and homology information
Function and homology information
 X-RAY DIFFRACTION /
X-RAY DIFFRACTION /  Authors
Authors Citation
Citation
 Structure visualization
Structure visualization Downloads & links
Downloads & links Other downloads
Other downloads
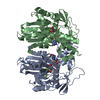
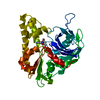

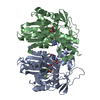
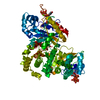
 PDBj
PDBj Assembly
Assembly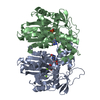



 Mass: 24.305 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: Mg
Mass: 24.305 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: Mg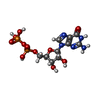
 Type: RNA linking / Mass: 443.201 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H15N5O11P2 / Comment: GDP, energy-carrying molecule*YM
Type: RNA linking / Mass: 443.201 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H15N5O11P2 / Comment: GDP, energy-carrying molecule*YM Mass: 18.015 Da / Num. of mol.: 522 / Source method: isolated from a natural source / Formula: H2O
Mass: 18.015 Da / Num. of mol.: 522 / Source method: isolated from a natural source / Formula: H2O Sample preparation
Sample preparation Processing
Processing