 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 2j0y | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | L-ficolin complexed to b-1,3-D-glucan | |||||||||
 Components Components | FICOLIN-2 | |||||||||
 Keywords Keywords | LECTIN / GLYCOPROTEIN / FICRINOGEN-LIKE / INNATE IMMUNITY / PATTERN RECOGNITION PROTEIN / COLLAGEN / IMMUNOLOGY / LECTIN- LIKE | |||||||||
| Function / homology |  Function and homology information Function and homology informationlipoteichoic acid immune receptor activity / mannan binding / recognition of apoptotic cell / Ficolins bind to repetitive carbohydrate structures on the target cell surface / Lectin pathway of complement activation / positive regulation of opsonization / collagen trimer / complement activation, lectin pathway / opsonization / cell surface pattern recognition receptor signaling pathway ...lipoteichoic acid immune receptor activity / mannan binding / recognition of apoptotic cell / Ficolins bind to repetitive carbohydrate structures on the target cell surface / Lectin pathway of complement activation / positive regulation of opsonization / collagen trimer / complement activation, lectin pathway / opsonization / cell surface pattern recognition receptor signaling pathway / carbohydrate derivative binding / symbiont cell surface / serine-type endopeptidase complex / proteoglycan binding / Initial triggering of complement / pattern recognition receptor activity / antigen binding / calcium-dependent protein binding / blood microparticle / defense response to Gram-negative bacterium / defense response to Gram-positive bacterium / external side of plasma membrane / signaling receptor binding / proteolysis / : / extracellular exosome / extracellular region / metal ion binding Similarity search - Function | |||||||||
| Biological species |  HOMO SAPIENS (human) HOMO SAPIENS (human) | |||||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / OTHER / Resolution: 2.35 Å X-RAY DIFFRACTION / SYNCHROTRON / OTHER / Resolution: 2.35 Å | |||||||||
 Authors Authors | Garlatti, V. / Gaboriaud, C. | |||||||||
 Citation Citation | Journal: Embo J. / Year: 2007 Title: Structural Insights Into the Innate Immune Recognition Specificities of L- and H-Ficolins. Authors: Garlatti, V. / Belloy, N. / Martin, L. / Lacroix, M. / Matsushita, M. / Endo, Y. / Fujita, T. / Fontecilla-Camps, J.C. / Arlaud, G.J. / Thielens, N.M. / Gaboriaud, C. | |||||||||
| History |
| |||||||||
| Remark 700 | SHEET THE SHEET STRUCTURE OF THIS MOLECULE IS BIFURCATED. IN ORDER TO REPRESENT THIS FEATURE IN ... SHEET THE SHEET STRUCTURE OF THIS MOLECULE IS BIFURCATED. IN ORDER TO REPRESENT THIS FEATURE IN THE SHEET RECORDS BELOW, TWO SHEETS ARE DEFINED. |
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 2j0y.cif.gz | 267.8 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb2j0y.ent.gz | 216.4 KB | Display | PDB format |
| PDBx/mmJSON format | 2j0y.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/j0/2j0yftp://data.pdbj.org/pub/pdb/validation_reports/j0/2j0y | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  2j0gC  2j0hC  2j1gC  2j2pC  2j3fC  2j3gC  2j3oC  2j3uC  2j5zC  2j60C  2j61C  2j64C C: citing same article ( |
|---|---|
| Similar structure data |

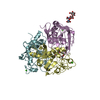
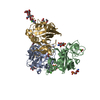
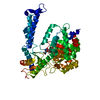

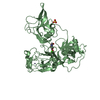

-Links
 PDBj
PDBj




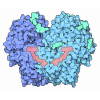
- Assembly
Assembly
| Deposited unit | 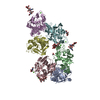
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2 | 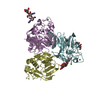
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Unit cell |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Noncrystallographic symmetry (NCS) | NCS domain:
NCS domain segments:
NCS ensembles :
|
-Components
-Protein , 1 types, 6 molecules ABCDEF
| #1: Protein | Mass: 24729.318 Da / Num. of mol.: 6 / Fragment: C-TERMINAL BINDING DOMAIN, RESIDUES 96-313 Source method: isolated from a genetically manipulated source Source: (gene. exp.) HOMO SAPIENS (human) / Tissue: PLASMA / Cell line (production host): High Five / Production host:   SPODOPTERA FRUGIPERDA (fall armyworm) / References: UniProt: Q15485 SPODOPTERA FRUGIPERDA (fall armyworm) / References: UniProt: Q15485 |
|---|
-Sugars , 4 types, 7 molecules 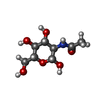
| #2: Polysaccharide | Source method: isolated from a genetically manipulated source #3: Polysaccharide | Source method: isolated from a genetically manipulated source #6: Sugar |  Type: D-saccharide, beta linking / Mass: 221.208 Da / Num. of mol.: 2 Type: D-saccharide, beta linking / Mass: 221.208 Da / Num. of mol.: 2Source method: isolated from a genetically manipulated source Formula: C8H15NO6 #7: Sugar | ChemComp-BGC / |  Type: D-saccharide, beta linking / Mass: 180.156 Da / Num. of mol.: 1 Type: D-saccharide, beta linking / Mass: 180.156 Da / Num. of mol.: 1Source method: isolated from a genetically manipulated source Formula: C6H12O6 |
|---|
-Non-polymers , 3 types, 301 molecules 


| #4: Chemical | ChemComp-ACT /  Mass: 59.044 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C2H3O2 Mass: 59.044 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C2H3O2 | ||
|---|---|---|---|
| #5: Chemical | ChemComp-CA /  Mass: 40.078 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: Ca Mass: 40.078 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: Ca#8: Water | ChemComp-HOH / | Mass: 18.015 Da / Num. of mol.: 294 / Source method: isolated from a natural source / Formula: H2O |
-Details
| Compound details | CALCIUM-DEPENDENT AND GLCNAC-BINDING LECTIN. |
|---|---|
| Has protein modification | Y |
| Sequence details | C-TERMINAL DOMAIN CONFLICTS BETWEEN UNP SEQUENCE AND COORDINATES: THERE IS A DIFFERENCE BECAUSE ...C-TERMINAL DOMAIN CONFLICTS BETWEEN UNP SEQUENCE AND COORDINATE |
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 2.52 Å3/Da / Density % sol: 50.8 % |
|---|
-Data collection
| Diffraction | Mean temperature: 100 K |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: ESRF  / Beamline: ID14-1 / Wavelength: 1.0718 / Beamline: ID14-1 / Wavelength: 1.0718 |
| Detector | Type: ADSC CCD / Detector: CCD / Date: Mar 19, 2006 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 1.0718 Å / Relative weight: 1 |
| Reflection | Resolution: 2.35→19.95 Å / Num. obs: 58888 / % possible obs: 98.1 % / Observed criterion σ(I): 0 / Redundancy: 4.6 % / Rmerge(I) obs: 0.09 / Net I/σ(I): 12 |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: OTHER / Resolution: 2.35→19.95 Å / Cor.coef. Fo:Fc: 0.948 / Cor.coef. Fo:Fc free: 0.915 / SU B: 16.678 / SU ML: 0.205 / TLS residual ADP flag: LIKELY RESIDUAL / Cross valid method: THROUGHOUT / ESU R: 0.405 / ESU R Free: 0.251 / Stereochemistry target values: MAXIMUM LIKELIHOOD Details: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS. THE A AND D CHAIN ARE NOT CLEARLY DEFINE IN DENSITY BECAUSE THEY ARE QUITE FLEXIBLE.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Ion probe radii: 0.8 Å / Shrinkage radii: 0.8 Å / VDW probe radii: 1.2 Å / Solvent model: BABINET MODEL WITH MASK | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 45.98 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 2.35→19.95 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
|