 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 1sjj | ||||||
|---|---|---|---|---|---|---|---|
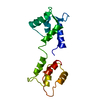
| Title | Cryo-EM Structure of Chicken Gizzard Smooth Muscle alpha-Actinin | ||||||
 Components Components | actinin | ||||||
 Keywords Keywords | CONTRACTILE PROTEIN / 3-helix bundle / calponin homology domain / calmodulin-like domain / actin binding protein | ||||||
| Function / homology |  Function and homology information Function and homology informationsmooth muscle dense body / Regulation of cytoskeletal remodeling and cell spreading by IPP complex components / RHOBTB2 GTPase cycle / : / Platelet degranulation / RHOF GTPase cycle / outer dense plaque of desmosome / inner dense plaque of desmosome / terminal web / zonula adherens ...smooth muscle dense body / Regulation of cytoskeletal remodeling and cell spreading by IPP complex components / RHOBTB2 GTPase cycle / : / Platelet degranulation / RHOF GTPase cycle / outer dense plaque of desmosome / inner dense plaque of desmosome / terminal web / zonula adherens / LIM domain binding / vinculin binding / muscle cell development / dense body / sarcomere organization / cortical actin cytoskeleton / cell leading edge / alpha-actinin binding / lateral plasma membrane / bicellular tight junction / skeletal muscle fiber development / ruffle / cell projection / stress fiber / phosphoprotein binding / sarcolemma / Z disc / actin filament binding / cell junction / lamellipodium / actin cytoskeleton organization / focal adhesion / calcium ion binding / protein homodimerization activity / plasma membrane Similarity search - Function | ||||||
| Biological species |  | ||||||
| Method | ELECTRON CRYSTALLOGRAPHY / electron crystallography / cryo EM / Resolution: 20 Å | ||||||
 Authors Authors | Liu, J. / Taylor, D.W. / Taylor, K.A. | ||||||
 Citation Citation | Journal: J Mol Biol / Year: 2004 Title: A 3-D reconstruction of smooth muscle alpha-actinin by CryoEm reveals two different conformations at the actin-binding region. Authors: Jun Liu / Dianne W Taylor / Kenneth A Taylor /  Abstract: Cryoelectron microscopy was used to obtain a 3-D image at 2.0 nm resolution of 2-D arrays of smooth muscle alpha-actinin. The reconstruction reveals a well-resolved long central domain with 90 ...Cryoelectron microscopy was used to obtain a 3-D image at 2.0 nm resolution of 2-D arrays of smooth muscle alpha-actinin. The reconstruction reveals a well-resolved long central domain with 90 degrees of left-handed twist and near 2-fold symmetry. However, the molecular ends which contain the actin binding and calmodulin-like domains, have different structures oriented approximately 90 degrees to each other. Atomic structures for the alpha-actinin domains were built by homology modeling and assembled into an atomic model. Model building suggests that in the 2-D arrays, the two calponin homology domains that comprise the actin-binding domain have a closed conformation at one end and an open conformation at the other end due to domain swapping. The open and closed conformations of the actin-binding domain suggests flexibility that may underlie Ca2+ regulation. The approximately 90 degrees orientation difference at the molecular ends may underlie alpha-actinin's ability to crosslink actin filaments in nearly any orientation. #1: Journal: J Mol Biol / Year: 1993 Title: Projection image of smooth muscle alpha-actinin from two-dimensional crystals formed on positively charged lipid layers. Authors: K A Taylor / D W Taylor / Abstract: Two-dimensional crystalline arrays of chicken gizzard alpha-actinin have been formed on positively charged lipid layers. This is the first reported crystallization of alpha-actinin. The crystals have ...Two-dimensional crystalline arrays of chicken gizzard alpha-actinin have been formed on positively charged lipid layers. This is the first reported crystallization of alpha-actinin. The crystals have unit cell dimensions of a = 248 A, b = 194 A, y = 106 degrees and contain two alpha-actinin molecules. The two-sided group is P21. Projection images obtained from electron micrographs of negatively stained crystals have been calculated to a resolution of 25 A. These images reveal a complex substructure. The molecule in projection is 340 A in length and has 12 density peaks that probably correspond to protein domains. A pair of peaks is found at each end of the molecule, these probably correspond to the actin binding region. Eight peaks are observed in the central, rod-shaped region, these may correspond to the spectrin-like repeats predicted from the amino acid sequence. However, these eight central peaks are not arranged in four pairs but, instead, consist of three central pairs flanked at either end by a single peak, which appears larger and denser in projection than the three central pairs. The individual alpha-actinin molecules in projection lack 2-fold symmetry suggesting that either smooth muscle alpha-actinin lacks a molecular 2-fold symmetry axis or that the molecular 2-fold is not parallel with the crystallographic 2-fold axis. The ends of the molecule have different appearance in projection, suggesting that the molecule is twisted about the long axis. A hypothesis is proposed to explain the variations in molecular length and Ca2+ sensitivity between alpha-actinin isoforms. #2: Journal: J Mol Biol / Year: 2001 Title: The three-dimensional structure of alpha-actinin obtained by cryoelectron microscopy suggests a model for Ca(2+)-dependent actin binding. Authors: J Tang / D W Taylor / K A Taylor / Abstract: The three-dimensional structure of alpha-actinin from rabbit skeletal muscle was determined by cryoelectron microscopy in combination with homology modeling of the separate domain structures based on ...The three-dimensional structure of alpha-actinin from rabbit skeletal muscle was determined by cryoelectron microscopy in combination with homology modeling of the separate domain structures based on results previously determined by X-ray crystallography and nuclear magnetic resonance spectroscopy. alpha-Actinin was induced to form two-dimensional arrays on a positively charged lipid monolayer and micrographs were collected from unstained, frozen hydrated specimens at tilt angles from 0 degrees to 60 degrees. Interpretation of the 15 A-resolution three-dimensional structure was done by manually docking homologous models of the three key domains, actin-binding, three-helix motif and the C-terminal calmodulin-like domains. The initial model was refined quantitatively to improve its fit to the experimental reconstruction. The molecular model of alpha-actinin provides the first view of the overall structure of a complete actin cross-linking protein. The structure is characterized by close proximity of the C-terminal, calmodulin-like domain to the linker between the two calponin-homology domains that comprise the actin-binding domain. This location suggests a hypothesis to explain the involvement of the C-terminal domain in Ca(2+)-dependent actin binding of non-muscle isoforms. #3: Journal: J Cell Biol / Year: 2000 Title: Isoforms of alpha-actinin from cardiac, smooth, and skeletal muscle form polar arrays of actin filaments. Authors: K A Taylor / D W Taylor / F Schachat / Abstract: We have used a positively charged lipid monolayer to form two-dimensional bundles of F-actin cross-linked by alpha-actinin to investigate the relative orientation of the actin filaments within them. ...We have used a positively charged lipid monolayer to form two-dimensional bundles of F-actin cross-linked by alpha-actinin to investigate the relative orientation of the actin filaments within them. This method prevents growth of the bundles perpendicular to the monolayer plane, thereby facilitating interpretation of the electron micrographs. Using alpha-actinin isoforms isolated from the three types of vertebrate muscle, i.e., cardiac, skeletal, and smooth, we have observed almost exclusively cross-linking between polar arrays of filaments, i.e., actin filaments with their plus ends oriented in the same direction. One type of bundle can be classified as an Archimedian spiral consisting of a single actin filament that spirals inward as the filament grows and the bundle is formed. These spirals have a consistent hand and grow to a limiting internal diameter of 0.4-0.7 microm, where the filaments appear to break and spiral formation ceases. These results, using isoforms usually characterized as cross-linkers of bipolar actin filament bundles, suggest that alpha-actinin is capable of cross-linking actin filaments in any orientation. Formation of specifically bipolar or polar filament arrays cross-linked by alpha-actinin may require additional factors that either determine the filament orientation or restrict the cross-linking capabilities of alpha-actinin. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 1sjj.cif.gz | 315.3 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb1sjj.ent.gz | 242.2 KB | Display | PDB format |
| PDBx/mmJSON format | 1sjj.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/sj/1sjjftp://data.pdbj.org/pub/pdb/validation_reports/sj/1sjj | HTTPS FTP |
|---|
-Related structure data
| Related structure data | |
|---|---|
| Similar structure data |








-Links
 PDBj
PDBj
- Assembly
Assembly
| Deposited unit | 
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||
| Unit cell |
|
-Components
| #1: Protein | Mass: 99873.156 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Details: Protein isolated from chicken gizzards / Source: (natural) |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON CRYSTALLOGRAPHY |
|---|---|
| EM experiment | Aggregation state: 2D ARRAY / 3D reconstruction method: electron crystallography |
- Sample preparation
Sample preparation
| Component | Name: alpha-actinin / Type: COMPLEX Details: The sample was formed on a positively charged lipid monolayer composed of dilauryl phosphatidyl choline and didodecyldimethyl ammonium bromide |
|---|---|
| Buffer solution | Name: Phosphate buffered KCl / pH: 7 / Details: Phosphate buffered KCl |
| Specimen | Conc.: 0.75 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Specimen support | Details: Samples were recovered from the lipid monolayer using reticulated carbon films, plunge frozen in liquid ethane, and stored for up to a month before use. |
| Vitrification | Details: Plunge frozen in liquid ethane |
| Crystal grow | *PLUS Method: electron crystallography / Details: electron crystallography |
-Data collection
| Microscopy | Model: FEI/PHILIPS CM300FEG/T |
|---|---|
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 24000 X / Nominal defocus max: 6000 nm / Nominal defocus min: 2000 nm / Cs: 2 mm |
| Specimen holder | Temperature: 100 K / Tilt angle max: 68 ° / Tilt angle min: 0 ° |
| Image recording | Film or detector model: KODAK SO-163 FILM Details: Developed in full strength D19 developer for 12 minutes. |
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Details: Correction of phases only | ||||||||||||||||||||||||||||
| 3D reconstruction | Method: 2-D crystallography / Resolution: 20 Å / Nominal pixel size: 5 Å / Actual pixel size: 5 Å Magnification calibration: Unit cell calibrated using the axial repeat of tobacco mosaic virus Details: Fourier transforms of images were merged using standard 2-D crystal programs with amplitudes and phases derived from images alone. Symmetry type: 2D CRYSTAL | ||||||||||||||||||||||||||||
| Atomic model building | Protocol: RIGID BODY FIT / Space: REAL Target criteria: highest cross correlation coefficient and lowest interatom conflicts Details: REFINEMENT PROTOCOL--rigid body | ||||||||||||||||||||||||||||
| Atomic model building |
| ||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST
| ||||||||||||||||||||||||||||
| Refinement | *PLUS Highest resolution: 20 Å | ||||||||||||||||||||||||||||
| Solvent computation | *PLUS | ||||||||||||||||||||||||||||
| Displacement parameters | *PLUS |
