 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
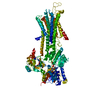
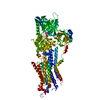
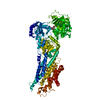
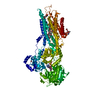
| 登録情報 | データベース: PDB / ID: 1kju | ||||||
|---|---|---|---|---|---|---|---|
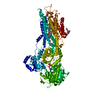
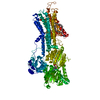
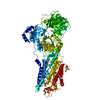
| タイトル | Ca2+-ATPase in the E2 State | ||||||
 要素 要素 | Sarcoplasmic/endoplasmic reticulum calcium ATPase 1a | ||||||
 キーワード キーワード | HYDROLASE / ION PUMP / CALCIUM / MEMBRANE PROTEIN / P-TYPE ATPASE / ACTIVE TRANSPORT / E2 / CRYO-EM | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報positive regulation of cardiac muscle cell contraction / positive regulation of calcium ion import into sarcoplasmic reticulum / positive regulation of ATPase-coupled calcium transmembrane transporter activity / positive regulation of fast-twitch skeletal muscle fiber contraction / H zone / regulation of striated muscle contraction / calcium ion import into sarcoplasmic reticulum / negative regulation of striated muscle contraction / P-type Ca2+ transporter / P-type calcium transporter activity ...positive regulation of cardiac muscle cell contraction / positive regulation of calcium ion import into sarcoplasmic reticulum / positive regulation of ATPase-coupled calcium transmembrane transporter activity / positive regulation of fast-twitch skeletal muscle fiber contraction / H zone / regulation of striated muscle contraction / calcium ion import into sarcoplasmic reticulum / negative regulation of striated muscle contraction / P-type Ca2+ transporter / P-type calcium transporter activity / I band / endoplasmic reticulum-Golgi intermediate compartment / sarcoplasmic reticulum membrane / sarcoplasmic reticulum / calcium ion transport / calcium ion binding / endoplasmic reticulum membrane / perinuclear region of cytoplasm / endoplasmic reticulum / ATP hydrolysis activity / ATP binding / membrane 類似検索 - 分子機能 | ||||||
| 生物種 |  | ||||||
| 手法 | 電子顕微鏡法 / らせん対称体再構成法 / クライオ電子顕微鏡法 / 解像度: 6 Å | ||||||
 データ登録者 データ登録者 | Xu, C. / Rice, W.J. / He, W. / Stokes, D.L. | ||||||
 引用 引用 | ジャーナル: J Mol Biol / 年: 2002 タイトル: A structural model for the catalytic cycle of Ca(2+)-ATPase. 著者: Chen Xu / William J Rice / Wanzhong He / David L Stokes /  要旨: Ca(2+)-ATPase is responsible for active transport of calcium ions across the sarcoplasmic reticulum membrane. This coupling involves an ordered sequence of reversible reactions occurring alternately ...Ca(2+)-ATPase is responsible for active transport of calcium ions across the sarcoplasmic reticulum membrane. This coupling involves an ordered sequence of reversible reactions occurring alternately at the ATP site within the cytoplasmic domains, or at the calcium transport sites within the transmembrane domain. These two sites are separated by a large distance and conformational changes have long been postulated to play an important role in their coordination. To characterize the nature of these conformational changes, we have built atomic models for two reaction intermediates and postulated the mechanisms governing the large structural changes. One model is based on fitting the X-ray crystallographic structure of Ca(2+)-ATPase in the E1 state to a new 6 A structure by cryoelectron microscopy in the E2 state. This fit indicates that calcium binding induces enormous movements of all three cytoplasmic domains as well as significant changes in several transmembrane helices. We found that fluorescein isothiocyanate displaced a decavanadate molecule normally located at the intersection of the three cytoplasmic domains, but did not affect their juxtaposition; this result indicates that our model likely reflects a native E2 conformation and not an artifact of decavanadate binding. To explain the dramatic structural effect of calcium binding, we propose that M4 and M5 transmembrane helices are responsive to calcium binding and directly induce rotation of the phosphorylation domain. Furthermore, we hypothesize that both the nucleotide-binding and beta-sheet domains are highly mobile and driven by Brownian motion to elicit phosphoenzyme formation and calcium transport, respectively. If so, the reaction cycle of Ca(2+)-ATPase would have elements of a Brownian ratchet, where the chemical reactions of ATP hydrolysis are used to direct the random thermal oscillations of an innately flexible molecule. #1: ジャーナル: Nature / 年: 2000タイトル: Crystal Structure of the Calcium Pump of Sarcoplasmic Reticulum at 2.6 A Resolution 著者: Toyoshima, C. / Nakasako, M. / Nomura, H. / Ogawa, H. #2: ジャーナル: Nature / 年: 1998タイトル: Structure of the Calcium Pump from Sarcoplasmic Reticulum at 8-A Resolution 著者: Zhang, P. / Toyoshima, C. / Yonekura, K. / Green, N.M. / Stokes, D.L. #3: ジャーナル: Ultramicroscopy / 年: 1997タイトル: Distortion Correction of Tubular Crystals: Improvements in the Acetylcholine Receptor Structure 著者: Beroukhim, R. / Unwin, N. #4: ジャーナル: Biophys.J. / 年: 2000タイトル: Modeling a Dehalogenase Fold into the 8-A Density Map for Ca(2+)-ATPase Defines a New Domain Structure 著者: Stokes, D.L. / Green, N.M. | ||||||
| 履歴 |
| ||||||
| Remark 999 | SEQUENCE THE SEQUENCE IN THIS ENTRY IS AN ISOFORM OF THE SEQUENCE IN SWISSPROT ENTRY P04191. THE C- ...SEQUENCE THE SEQUENCE IN THIS ENTRY IS AN ISOFORM OF THE SEQUENCE IN SWISSPROT ENTRY P04191. THE C-TERMINAL REGION OF P04191 CONSISTS OF RESIDUES 994-1001, DPEDERRK. IN THIS ENTRY, THE C-TERMINAL RESIDUE IS 994, G. |
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 1kju.cif.gz | 200.2 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb1kju.ent.gz | 159.1 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 1kju.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 1kju_validation.pdf.gz | 365.2 KB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 1kju_full_validation.pdf.gz | 401.5 KB | 表示 | |
| XML形式データ | 1kju_validation.xml.gz | 23.4 KB | 表示 | |
| CIF形式データ | 1kju_validation.cif.gz | 34.2 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/kj/1kjuftp://data.pdbj.org/pub/pdb/validation_reports/kj/1kju | HTTPS FTP |
-関連構造データ
| 関連構造データ | |
|---|---|
| 類似構造データ |




-リンク
 PDBj
PDBj
- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
| #1: タンパク質 | 分子量: 109602.578 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: HELICAL ARRAY / 3次元再構成法: らせん対称体再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Ca2+-ATPase tubular crystals / タイプ: COMPLEX 詳細: Crystals formed in presence of decavanadate and thapsigargin | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 緩衝液 | 名称: 20 mM imidazole pH 7.4, 100 mM KCl, 5 mM MgCl2, 0.5 mM EGTA, 0.5 mM decavanadate, 0.01 mM thapsigargin pH: 7.4 詳細: 20 mM imidazole pH 7.4, 100 mM KCl, 5 mM MgCl2, 0.5 mM EGTA, 0.5 mM decavanadate, 0.01 mM thapsigargin | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 試料 | 濃度: 0.05 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 急速凍結 | 詳細: Samples at approximately 1 mg/ml were diluted 1/20 immediately prior to application to glow-discharged fenestrated carbon grids. All freezing was done in cold room. After blotting, samples ...詳細: Samples at approximately 1 mg/ml were diluted 1/20 immediately prior to application to glow-discharged fenestrated carbon grids. All freezing was done in cold room. After blotting, samples were plunged into liquid ethane for freezing and stored in liquid nitrogen. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 結晶化 | *PLUS 温度: 4 ℃ / 手法: microdialysis | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶液の組成 | *PLUS
|
- 電子顕微鏡撮影
電子顕微鏡撮影
| 顕微鏡 | モデル: FEI/PHILIPS CM200FEG / 日付: 2000年1月1日 |
|---|---|
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 200 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 200 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 50000 X / 倍率(補正後): 51300 X / 最大 デフォーカス(公称値): 1400 nm / 最小 デフォーカス(公称値): 700 nm / Cs: 2 mm |
| 試料ホルダ | 温度: 100 K / 傾斜角・最大: 0 ° / 傾斜角・最小: 0 ° |
| 撮影 | 電子線照射量: 10 e/Å2 / フィルム・検出器のモデル: KODAK SO-163 FILM |
| 画像スキャン | デジタル画像の数: 58 |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | 詳細: CTF correction of each particle with 4.6% amplitude contrast | ||||||||||||||||||||
| 3次元再構成 | 手法: helical reconstruction / 解像度: 6 Å / 粒子像の数: 95 / ピクセルサイズ(実測値): 2.53 Å / 倍率補正: light harvesting complex crystals 詳細: Combination of Fourier and real space averaging. Helical Reconstruction: A total of 95 repeats from 58 individual tubes were used for the reconstruction. These tubes fell into 5 helical ...詳細: Combination of Fourier and real space averaging. Helical Reconstruction: A total of 95 repeats from 58 individual tubes were used for the reconstruction. These tubes fell into 5 helical symmetries, defined by the Bessel order (n) of the principal layer lines (1,0) and (0,1). These 5 symmetries were (-23,6), (-22,6), (-21,6), (-20,6) and (-19,6). For the reference symmetry (-22,6), the unit cell parameters were: a= 56.9A, b=117.1A, gamma=64.2 deg. Data within each helical symmetry were averaged in Fourier space and distortion-correction techniques were applied (Beroukhim and Unwin, 1997). For each repeat, unit cell parameters were calculated, and repeats differing by more than 1.5% of the average value were discarded. The CTF was used to correct phases and weight amplitudes prior to averaging within each helical symmetry. Finally, maps were calculated from each of the averaged datasets. Each of the 2 molecules composing the unit cell were masked and aligned with the corresponding molecule from the reference map (-22,6). The maps were averaged in real-space then back-transformed into Fourier space. Two-fold symmetry was constrained before calculating the final map. Effective resolution of the reconstruction: The resolution of the final reconstruction was determined to be 6 A by two methods. First, the dataset was split into two equal parts and two independant reconstructions were made. After masking and aligning molecules from these maps, Fourier shell correlation coefficients and associated phase residuals were calculated. Secondly, since the crystals had p2 symmetry, two-fold phase residuals of layer lined datasets were used to monitor resolution. 対称性のタイプ: HELICAL | ||||||||||||||||||||
| 原子モデル構築 | 空間: REAL 詳細: DETAILS--The atomic coordinates of Ca-ATPase (1EUL) were divided into 4 domains by breaking the peptide at 3 hinge points. Each domain was separately fit into the electron density map by ...詳細: DETAILS--The atomic coordinates of Ca-ATPase (1EUL) were divided into 4 domains by breaking the peptide at 3 hinge points. Each domain was separately fit into the electron density map by manual docking using the program O. Besides matching overall shape, the major criteria involved matching a-helices into strong columns of density in the map. Whereas cytoplasmic domains were treated as rigid bodies, elements of the transmembrane domain were bent or displaced to match strong densities in the cryo-em map. Finally, the loops composing the 3 hinges were rebuilt to reconnect the domains and X-PLOR was used to perform local energy minimization. | ||||||||||||||||||||
| 原子モデル構築 | PDB-ID: 1EUL 1eul Accession code: 1EUL / Source name: PDB / タイプ: experimental model | ||||||||||||||||||||
| 精密化 | 最高解像度: 6 Å | ||||||||||||||||||||
| 精密化ステップ | サイクル: LAST / 最高解像度: 6 Å
| ||||||||||||||||||||
| ソフトウェア | *PLUS 名称: X-PLOR / 分類: refinement | ||||||||||||||||||||
| 拘束条件 | *PLUS
|
