 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 1hb9 | ||||||
|---|---|---|---|---|---|---|---|
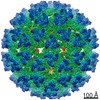
| タイトル | quasi-atomic resolution model of bacteriophage PRD1 wild type virion, obtained by combined cryo-EM and X-ray crystallography. | ||||||
 要素 要素 | BACTERIOPHAGE PRD1 | ||||||
 キーワード キーワード | VIRUS / VIRUS/VIRAL PROTEIN / TECTIVIRIDAE / BACTERIOPHAGE PRD1 / CRYO- EM / IMAGE RECONSTRUCTION / ICOSAHEDRAL VIRUS | ||||||
| 機能・相同性 | Bacteriophage PRD1, P3 / Bacteriophage PRD1, P3, N-terminal / P3 major capsid protein N-terminal / Group II dsDNA virus coat/capsid protein / Viral coat protein subunit / viral capsid / Major capsid protein P3 機能・相同性情報 機能・相同性情報 | ||||||
| 生物種 |   BACTERIOPHAGE PRD1 (ファージ) BACTERIOPHAGE PRD1 (ファージ) | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 25 Å | ||||||
 データ登録者 データ登録者 | San Martin, C. / Burnett, R.M. / De Haas, F. / Heinkel, R. / Rutten, T. / Fuller, S.D. / Butcher, S.J. / Bamford, D.H. | ||||||
 引用 引用 | ジャーナル: Structure / 年: 2001 タイトル: Combined EM/X-ray imaging yields a quasi-atomic model of the adenovirus-related bacteriophage PRD1 and shows key capsid and membrane interactions. 著者: C S Martín / R M Burnett / F de Haas / R Heinkel / T Rutten / S D Fuller / S J Butcher / D H Bamford /  要旨: BACKGROUND: The dsDNA bacteriophage PRD1 has a membrane inside its icosahedral capsid. While its large size (66 MDa) hinders the study of the complete virion at atomic resolution, a 1.65-A ...BACKGROUND: The dsDNA bacteriophage PRD1 has a membrane inside its icosahedral capsid. While its large size (66 MDa) hinders the study of the complete virion at atomic resolution, a 1.65-A crystallographic structure of its major coat protein, P3, is available. Cryo-electron microscopy (cryo-EM) and three-dimensional reconstruction have shown the capsid at 20-28 A resolution. Striking architectural similarities between PRD1 and the mammalian adenovirus indicate a common ancestor. RESULTS: The P3 atomic structure has been fitted into improved cryo-EM reconstructions for three types of PRD1 particles: the wild-type virion, a packaging mutant without DNA, and a P3-shell lacking ...RESULTS: The P3 atomic structure has been fitted into improved cryo-EM reconstructions for three types of PRD1 particles: the wild-type virion, a packaging mutant without DNA, and a P3-shell lacking the membrane and the vertices. Establishing the absolute EM scale was crucial for an accurate match. The resulting "quasi-atomic" models of the capsid define the residues involved in the major P3 interactions, within the quasi-equivalent interfaces and with the membrane, and show how these are altered upon DNA packaging. CONCLUSIONS: The new cryo-EM reconstructions reveal the structure of the PRD1 vertex and the concentric packing of DNA. The capsid is essentially unchanged upon DNA packaging, with alterations ...CONCLUSIONS: The new cryo-EM reconstructions reveal the structure of the PRD1 vertex and the concentric packing of DNA. The capsid is essentially unchanged upon DNA packaging, with alterations limited to those P3 residues involved in membrane contacts. These are restricted to a few of the N termini along the icosahedral edges in the empty particle; DNA packaging leads to a 4-fold increase in the number of contacts, including almost all copies of the N terminus and the loop between the two beta barrels. Analysis of the P3 residues in each quasi-equivalent interface suggests two sites for minor proteins in the capsid edges, analogous to those in adenovirus. #1: ジャーナル: Acta Crystallogr D Biol Crystallogr / 年: 2002タイトル: The X-ray crystal structure of P3, the major coat protein of the lipid-containing bacteriophage PRD1, at 1.65 A resolution. 著者: Stacy D Benson / Jaana K H Bamford / Dennis H Bamford / Roger M Burnett / 要旨: P3 has been imaged with X-ray crystallography to reveal a trimeric molecule with strikingly similar characteristics to hexon, the major coat protein of adenovirus. The structure of native P3 has now ...P3 has been imaged with X-ray crystallography to reveal a trimeric molecule with strikingly similar characteristics to hexon, the major coat protein of adenovirus. The structure of native P3 has now been extended to 1.65 A resolution (R(work) = 19.0% and R(free) = 20.8%). The new high-resolution model shows that P3 forms crystals through hydrophobic patches solvated by 2-methyl-2,4-pentanediol molecules. It reveals details of how the molecule's high stability may be achieved through ordered solvent in addition to intra- and intersubunit interactions. Of particular importance is a 'puddle' at the top of the molecule containing a four-layer deep hydration shell that cross-links a complex structural feature formed by 'trimerization loops'. These loops also link subunits by extending over a neighbor to reach the third subunit in the trimer. As each subunit has two eight-stranded viral jelly rolls, the trimer has a pseudo-hexagonal shape to allow close packing in its 240 hexavalent capsid positions. Flexible regions in P3 facilitate these interactions within the capsid and with the underlying membrane. A selenometh-ionine P3 derivative, with which the structure was solved, has been refined to 2.2 A resolution (R(work) = 20.1% and R(free) = 22.8%). The derivatized molecule is essentially unchanged, although synchrotron radiation has the curious effect of causing it to rotate about its threefold axis. P3 is a second example of a trimeric 'double-barrel' protein that forms a stable building block with optimal shape for constructing a large icosahedral viral capsid. A major difference is that hexon has long variable loops that distinguish different adenovirus species. The short loops in P3 and the severe constraints of its various interactions explain why the PRD1 family has highly conserved coat proteins. #2: ジャーナル: Cell / 年: 1999タイトル: Viral evolution revealed by bacteriophage PRD1 and human adenovirus coat protein structures. 著者: S D Benson / J K Bamford / D H Bamford / R M Burnett / 要旨: The unusual bacteriophage PRD1 features a membrane beneath its icosahedral protein coat. The crystal structure of the major coat protein, P3, at 1.85 A resolution reveals a molecule with three ...The unusual bacteriophage PRD1 features a membrane beneath its icosahedral protein coat. The crystal structure of the major coat protein, P3, at 1.85 A resolution reveals a molecule with three interlocking subunits, each with two eight-stranded viral jelly rolls normal to the viral capsid, and putative membrane-interacting regions. Surprisingly, the P3 molecule closely resembles hexon, the equivalent protein in human adenovirus. Both viruses also have similar overall architecture, with identical capsid lattices and attachment proteins at their vertices. Although these two dsDNA viruses infect hosts from very different kingdoms, their striking similarities, from major coat protein through capsid architecture, strongly suggest their evolutionary relationship. #3: ジャーナル: EMBO J / 年: 1995 タイトル: DNA packaging orders the membrane of bacteriophage PRD1. 著者: S J Butcher / D H Bamford / S D Fuller /  要旨: Bacteriophage PRD1 contains a linear dsDNA genome enclosed by a lipid membrane lying within a protein coat. Determination of the structure of the detergent-treated particle to 2 nm by cryo-electron ...Bacteriophage PRD1 contains a linear dsDNA genome enclosed by a lipid membrane lying within a protein coat. Determination of the structure of the detergent-treated particle to 2 nm by cryo-electron microscopy and three-dimensional reconstruction has defined the position of the major coat protein P3. The coat contains 240 copies of trimeric P3 packed into positions of local 6-fold symmetry on a T = 25 lattice. The three-dimensional structures of the PRD1 virion and a DNA packaging mutant to a resolution of 2.8 nm have revealed specific interactions between the coat and the underlying membrane. The membrane is clearly visible as two leaflets separated by 2 nm and spanned by transmembrane density. The size of the coat does not change upon DNA packaging. Instead, the number of interactions seen between the protein shell and the membrane and the order of the membrane components increase. Thus the membrane of PRD1 plays a role in assembly which is akin to that played by the nucleocapsid in other membrane viruses. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 1hb9.cif.gz | 861.1 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb1hb9.ent.gz | 725.8 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 1hb9.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 1hb9_validation.pdf.gz | 1.1 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 1hb9_full_validation.pdf.gz | 1.1 MB | 表示 | |
| XML形式データ | 1hb9_validation.xml.gz | 121 KB | 表示 | |
| CIF形式データ | 1hb9_validation.cif.gz | 183.4 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/hb/1hb9ftp://data.pdbj.org/pub/pdb/validation_reports/hb/1hb9 | HTTPS FTP |
-関連構造データ
















-リンク
 PDBj
PDBj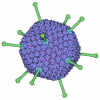

- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 | x 60
|
| 2 |
|
| 3 | x 5
|
| 4 | x 6
|
| 5 | 
|
| 対称性 | 点対称性: (シェーンフリース記号: I (正20面体型対称)) |
-要素
| #1: タンパク質 | 分子量: 43346.219 Da / 分子数: 12 / 由来タイプ: 組換発現 / 由来: (組換発現) BACTERIOPHAGE PRD1 (ファージ) / 株: DS88 / 発現宿主:  SALMONELLA TYPHIMURIUM (サルモネラ菌) / 株 (発現宿主): DS88 / 参照: UniProt: P22535 SALMONELLA TYPHIMURIUM (サルモネラ菌) / 株 (発現宿主): DS88 / 参照: UniProt: P22535 |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: BACTERIOPHAGE PRD1 / タイプ: VIRUS 詳細: 400 MESH COPPER GLOW DISCHARGE SAMPLES WERE PREPARED AS THIN LAYERS OF VITREOUS ICE |
|---|---|
| 緩衝液 | pH: 7.2 |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 試料支持 | 詳細: HOLEY CARBON |
| 急速凍結 | 装置: HOMEMADE PLUNGER / 凍結剤: ETHANE / 詳細: PLUNGE VITRIFICATION |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 顕微鏡 | モデル: FEI/PHILIPS CM200FEG / 日付: 1998年6月15日 詳細: SAMPLES WERE MAINTAINED AT LIQUID NITROGEN TEMPERATURES IN THE MICROSCOPE WITH A GATAN 626-0300 CRYOTRANSFER HOLDER |
|---|---|
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 200 kV / 照射モード: OTHER FIELD EMISSION GUN / 加速電圧: 200 kV / 照射モード: OTHER |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 36000 X / 最大 デフォーカス(公称値): 4100 nm / 最小 デフォーカス(公称値): 1300 nm / Cs: 2 mm |
| 試料ホルダ | 温度: 95 K / 傾斜角・最大: 0 ° / 傾斜角・最小: 0 ° |
| 撮影 | 電子線照射量: 10 e/Å2 / フィルム・検出器のモデル: KODAK SO-163 FILM |
| 画像スキャン | デジタル画像の数: 12 |
| 放射波長 | 相対比: 1 |
- 解析
解析
| EMソフトウェア |
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 対称性 | 点対称性: I (正20面体型対称) | ||||||||||||
| 3次元再構成 | 手法: ICOSAHEDRAL / 解像度: 25 Å / 粒子像の数: 1800 / ピクセルサイズ(公称値): 3.68 Å / ピクセルサイズ(実測値): 3.42 Å 倍率補正: THE PIXEL SIZE OF THE CRYO-EM MAP WAS OBTAINED USING THE X-RAY STRUCTURE OF THE P3 TRIMER AS A REFERENCE. AFTER AN INITIAL FITTING USING THE NOMINAL PIXEL SIZE, THE P3 TRIMERS IN THE ...倍率補正: THE PIXEL SIZE OF THE CRYO-EM MAP WAS OBTAINED USING THE X-RAY STRUCTURE OF THE P3 TRIMER AS A REFERENCE. AFTER AN INITIAL FITTING USING THE NOMINAL PIXEL SIZE, THE P3 TRIMERS IN THE ICOSAHEDRAL ASYMMETRIC UNIT WERE GRADUALLY TRANSLATED TOWARDS THE CENTER OF THE PARTICLE UNTIL THE CRYSTALLOGRAPHIC R-FACTOR WAS MINIMISED. 詳細: THE ORIENTATIONS WERE REFINED BY THE CROSS COMMON LINES LINES METHOD (SIMPLEX) AND THE POLAR FOURIER TRANSFORM METHOD. MODEL-BASED, POLAR-FOURIER-TRANSFORM (FULLER ET AL. 1996, J.STRUC.BIOL. ...詳細: THE ORIENTATIONS WERE REFINED BY THE CROSS COMMON LINES LINES METHOD (SIMPLEX) AND THE POLAR FOURIER TRANSFORM METHOD. MODEL-BASED, POLAR-FOURIER-TRANSFORM (FULLER ET AL. 1996, J.STRUC.BIOL. 116, 48-55; BAKER AND CHENG, 1996, J.STRUC.BIOL. 116, 120-130) MODEL-BASED CROSS COMMON LINES SEARCH AND REFINEMENT (CROWTHER ET AL. 1970, NATURE (LONDON) 226, 421-425; FULLER ET AL. 1996, J.STRUC.BIOL. 116, 48-55; FERLENGHI ET AL. 1998, J.MOL.BIOL. 283, 71-81). THE EFFECTIVE RESOLUTION OF THE FINAL RECONSTRUCTED DENSITY WAS DETERMINED TO BE AT LEAST 25 ANGSTROMS, AS MEASURED BY RANDOMLY SPLITTING THE PARTICLES INTO TWO SETS AND CALCULATING THE FOURIER SHELL CORRELATION OBTAINED FROM SEPARATE RECONSTRUCTIONS (HARAUZ AND VAN HEEL 1986, OPTIK 73, 146-156). THE EIGENVALUE SPECTRUM GAVE AN INDICATION OF THE RANDOMNESS OF THE DATA THAT WAS INCLUDED IN THE RECONSTRUCTION. THE COMPLETENESS OF THE DATA WAS VERIFIED IN THAT ALL EIGENVALUES EXCEEDED 100. THE COORDINATES ARE IN THE P, Q, R FRAME IN ANGSTROM UNITS AND CORRESPOND TO ICOSAHEDRAL SYMMETRY AXES. THE ORIGIN IS CHOSEN AT THE CENTER OF THE VIRUS WITH P, Q AND R ALONG MUTUALLY PERPENDICULAR TWO-FOLD AXES OF THE ICOSAHEDRON. THEY SHOULD REMAIN IN THAT FRAME FOR THE EASE OF THE USER IN CREATING THE BIOLOGICALLY SIGNIFICANT VIRAL COMPLEX PARTICLE USING THE 60 ICOSAHEDRAL SYMMETRY OPERATORS. RESIDUES NOT VISIBLE IN THE ORIGINAL CRYSTAL STRUCTURES ARE NOT INCLUDED IN THE CRYO-EM STRUCTURE MODEL. 対称性のタイプ: POINT | ||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT / 空間: RECIPROCAL / Target criteria: R-factor 詳細: METHOD--THE CRYSTAL STRUCTURE OF THE MAJOR COAT PROTEIN P3 (PDB FILE 1HX6) WAS PLACED INTO THE CRYO-EM DENSITY MAP. THE CAPSID PROTEIN WAS FIRST MANUALLY POSITIONED INTO THE CRYO-EM DENSITY ...詳細: METHOD--THE CRYSTAL STRUCTURE OF THE MAJOR COAT PROTEIN P3 (PDB FILE 1HX6) WAS PLACED INTO THE CRYO-EM DENSITY MAP. THE CAPSID PROTEIN WAS FIRST MANUALLY POSITIONED INTO THE CRYO-EM DENSITY CORRESPONDING TO POSITIONS OF THE FOUR INDEPENDENT TRIMERS IN THE ICOSAHEDRAL ASYMMETRIC UNIT. THESE POSITIONS WERE THEN REFINED BY RIGID BODY REFINEMENT IN RECIPROCAL SPACE WITH THE PROGRAM XPLOR. QUALITY OF THE FIT R- FACTOR= 0.360, CROSS-CORRELATI0N COEFFICIENT 0.880, ATOMS OUTSIDE DENSITY PER ICOSAHEDRAL ASYMMETRIC UNIT 801 (2.3%), ATOM CLASHES PER ICOSAHEDRAL ASYMMETRIC UNIT 69 (0.2%) REFINEMENT PROTOCOL--RIGID BODY REFINEMENT | ||||||||||||
| 原子モデル構築 | PDB-ID: 1HX6 Accession code: 1HX6 / Source name: PDB / タイプ: experimental model | ||||||||||||
| 精密化 | 最高解像度: 25 Å | ||||||||||||
| 精密化ステップ | サイクル: LAST / 最高解像度: 25 Å
|
