 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | 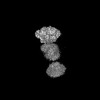 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | CryoEM structure of SV40 LTag bound to SV40 origin DNA | |||||||||
 Map data Map data | ||||||||||
 Sample Sample |
| |||||||||
 Keywords Keywords | Helicase / ATPase / SV40 origin DNA / AMP-PNP / DNA replication / TRANSLOCASE | |||||||||
| Biological species |  Betapolyomavirus macacae Betapolyomavirus macacae | |||||||||
| Method | single particle reconstruction / cryo EM / Resolution: 3.7 Å | |||||||||
 Authors Authors | Danazumi AU / Shahid T / Tehseen M / Alhudhali L / Clark A / Savva CG / Hamdan SM / De Biasio A | |||||||||
| Funding support |  Saudi Arabia, 1 items Saudi Arabia, 1 items
| |||||||||
 Citation Citation | Journal: Nature / Year: 2025 Title: Structural dynamics of DNA unwinding by a replicative helicase. Authors: Taha Shahid / Ammar U Danazumi / Muhammad Tehseen / Lubna Alhudhali / Alice R Clark / Christos G Savva / Samir M Hamdan / Alfredo De Biasio /  Abstract: Hexameric helicases are nucleotide-driven molecular machines that unwind DNA to initiate replication across all domains of life. Despite decades of intensive study, several critical aspects of their ...Hexameric helicases are nucleotide-driven molecular machines that unwind DNA to initiate replication across all domains of life. Despite decades of intensive study, several critical aspects of their function remain unresolved: the site and mechanism of DNA strand separation, the mechanics of unwinding propagation, and the dynamic relationship between nucleotide hydrolysis and DNA movement. Here, using cryo-electron microscopy (cryo-EM), we show that the simian virus 40 large tumour antigen (LTag) helicase assembles in the form of head-to-head hexamers at replication origins, melting DNA at two symmetrically positioned sites to establish bidirectional replication forks. Through continuous heterogeneity analysis, we characterize the conformational landscape of LTag on forked DNA under catalytic conditions, demonstrating coordinated motions that drive DNA translocation and unwinding. We show that the helicase pulls the tracking strand through DNA-binding loops lining the central channel, while directing the non-tracking strand out of the rear, in a cyclic process. ATP hydrolysis functions as an 'entropy switch', removing blocks to translocation rather than directly powering DNA movement. Our structures show the allosteric couplings between nucleotide turnover and subunit motions that enable DNA unwinding while maintaining dedicated exit paths for the separated strands. These findings provide a comprehensive model for replication fork establishment and progression that extends from viral to eukaryotic systems. More broadly, they introduce fundamental principles of the mechanism by which ATP-dependent enzymes achieve efficient mechanical work through entropy-driven allostery. | |||||||||
| History |
|
- Structure visualization
Structure visualization
- Downloads & links
Downloads & links
-EMDB archive
| Map data | emd_62502.map.gz | 513.4 MB |  EMDB map data format EMDB map data format | |
|---|---|---|---|---|
| Header (meta data) | emd-62502-v30.xmlemd-62502.xml | 14.7 KB 14.7 KB | Display Display | EMDB header |
| FSC (resolution estimation) | emd_62502_fsc.xml | 19.1 KB | Display | FSC data file |
| Images |  emd_62502.png emd_62502.png | 61.7 KB | ||
| Masks | emd_62502_msk_1.map | 600.7 MB | Mask map | |
| Filedesc metadata | emd-62502.cif.gz | 5.2 KB | ||
| Others | emd_62502_half_map_1.map.gzemd_62502_half_map_2.map.gz | 504.1 MB 504.4 MB | ||
| Archive directory |  http://ftp.pdbj.org/pub/emdb/structures/EMD-62502ftp://ftp.pdbj.org/pub/emdb/structures/EMD-62502 http://ftp.pdbj.org/pub/emdb/structures/EMD-62502ftp://ftp.pdbj.org/pub/emdb/structures/EMD-62502 | HTTPS FTP |
-Validation report
| Summary document | emd_62502_validation.pdf.gz | 1020 KB | Display | EMDB validaton report |
|---|---|---|---|---|
| Full document | emd_62502_full_validation.pdf.gz | 1019.6 KB | Display | |
| Data in XML | emd_62502_validation.xml.gz | 25.8 KB | Display | |
| Data in CIF | emd_62502_validation.cif.gz | 34.3 KB | Display | |
| Arichive directory | https://ftp.pdbj.org/pub/emdb/validation_reports/EMD-62502ftp://ftp.pdbj.org/pub/emdb/validation_reports/EMD-62502 | HTTPS FTP |
-Related structure data
| Related structure data |  9evhC  9evpC  9exdC  9f3tC  9f3uC  9f5iC  9f73C  9f74C  9f75C  9f7nC  9f9nC  9f9oC  9f9wC  9f9xC  9fa1C  9fa2C  9fb0C  9fb4C  9fb5C  9fb6C  9kaeC  9kakC C: citing same article ( |
|---|
































-Links
| EMDB pages | EMDB (EBI/PDBe) / EMDataResource |
|---|
-Map
| File | Download / File: emd_62502.map.gz / Format: CCP4 / Size: 600.7 MB / Type: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Voxel size | X=Y=Z: 0.93 Å | ||||||||||||||||||||
| Density |
| ||||||||||||||||||||
| Symmetry | Space group: 1 | ||||||||||||||||||||
| Details | EMDB XML:
|
-Supplemental data
- Sample components
Sample components
-Entire : Complex of SV40 LTag with SV40 origin DNA
| Entire | Name: Complex of SV40 LTag with SV40 origin DNA |
|---|---|
| Components |
|
-Supramolecule #1: Complex of SV40 LTag with SV40 origin DNA
| Supramolecule | Name: Complex of SV40 LTag with SV40 origin DNA / type: complex / ID: 1 / Parent: 0 / Macromolecule list: all |
|---|---|
| Source (natural) | Organism: Betapolyomavirus macacae |
| Molecular weight | Theoretical: 1 MDa |
-Macromolecule #1: LTag
| Macromolecule | Name: LTag / type: other / ID: 1 / Classification: other |
|---|---|
| Source (natural) | Organism: Betapolyomavirus macacae |
| Sequence | String: MDKVLNREES LQLMDLLGLE RSAWGNIPLM RKAYLKKCKE FHPDKGGDEE KMKKMNTLYK KMEDGVKYA HQPDFGGFWD ATEIPTYGTD EWEQWWNAFN EENLFCSEEM PSSDDEATAD S QHSTPPKK KRKVEDPKDF PSELLSFLSH AVFSNRTLAC FAIYTTKEKA ...String: MDKVLNREES LQLMDLLGLE RSAWGNIPLM RKAYLKKCKE FHPDKGGDEE KMKKMNTLYK KMEDGVKYA HQPDFGGFWD ATEIPTYGTD EWEQWWNAFN EENLFCSEEM PSSDDEATAD S QHSTPPKK KRKVEDPKDF PSELLSFLSH AVFSNRTLAC FAIYTTKEKA ALLYKKIMEK YS VTFISRH NSYNHNILFF LTPHRHRVSA INNYAQKLCT FSFLICKGVN KEYLMYSALT RDP FSVIEE SLPGGLKEHD FNPEEAEETK QVSWKLVTEY AMETKCDDVL LLLGMYLEFQ YSFE MCLKC IKKEQPSHYK YHEKHYANAA IFADSKNQKT ICQQAVDTVL AKKRVDSLQL TREQM LTNR FNDLLDRMDI MFGSTGSADI EEWMAGVAWL HCLLPKMDSV VYDFLKCMVY NIPKKR YWL FKGPIDSGKT TLAAALLELC GGKALNVNLP LDRLNFELGV AIDQFLVVFE DVKGTGG ES RDLPSGQGIN NLDNLRDYLD GSVKVNLEKK HLNKRTQIFP PGIVTMNEYS VPKTLQAR F VKQIDFRPKD YLKHCLERSE FLLEKRIIQS GIALLLMLIW YRPVAEFAQS IQSRIVEWK ERLDKEFSLS VYQKMKFNVA MGIGVLDWLR NSDDDDEDSQ ENADKNEDGG EKNMEDSGHE TGIDSQSQG SFQAPQSSQS VHDHNQPYHI CRGFTCFKKP PTPPPEPET |
| Recombinant expression | Organism:   Spodoptera frugiperda (fall armyworm) Spodoptera frugiperda (fall armyworm) |
-Experimental details
-Structure determination
| Method | cryo EM |
|---|---|
 Processing Processing | single particle reconstruction |
| Aggregation state | particle |
-Sample preparation
| Concentration | 0.5 mg/mL |
|---|---|
| Buffer | pH: 8 |
| Vitrification | Cryogen name: ETHANE / Chamber humidity: 100 % / Chamber temperature: 277.15 K / Instrument: FEI VITROBOT MARK IV |
- Electron microscopy
Electron microscopy
| Microscope | TFS KRIOS |
|---|---|
| Image recording | Film or detector model: FEI FALCON IV (4k x 4k) / Average exposure time: 5.0 sec. / Average electron dose: 41.28 e/Å2 |
| Electron beam | Acceleration voltage: 300 kV / Electron source:  FIELD EMISSION GUN FIELD EMISSION GUN |
| Electron optics | C2 aperture diameter: 50.0 µm / Illumination mode: FLOOD BEAM / Imaging mode: BRIGHT FIELD / Cs: 2.7 mm / Nominal defocus max: 2.6 µm / Nominal defocus min: 1.1 µm / Nominal magnification: 130000 |
| Sample stage | Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER / Cooling holder cryogen: NITROGEN |
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
+Image processing
-Atomic model buiding 1
| Refinement | Protocol: FLEXIBLE FIT |
|---|
