Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- EMDB-49510: Structure of substrates-engaged MIDN-bound human 26S proteasome,E... -
+ Open data
Open data

- Basic information
Basic information
| Entry |  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
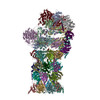
| Title | Structure of substrates-engaged MIDN-bound human 26S proteasome,ED-MIDN state (Composite map) | |||||||||
 Map data Map data | ||||||||||
 Sample Sample |
| |||||||||
 Keywords Keywords | Ubiquitin-indepedent / proteasome / Midnolin / 26S complex / ED-state / IMMUNE SYSTEM | |||||||||
| Function / homology |  Function and homology information Function and homology informationthyrotropin-releasing hormone receptor binding / nuclear proteasome complex / host-mediated perturbation of viral transcription / positive regulation of inclusion body assembly / Impaired BRCA2 translocation to the nucleus / Impaired BRCA2 binding to SEM1 (DSS1) / meiosis I / proteasome accessory complex / purine ribonucleoside triphosphate binding / integrator complex ...thyrotropin-releasing hormone receptor binding / nuclear proteasome complex / host-mediated perturbation of viral transcription / positive regulation of inclusion body assembly / Impaired BRCA2 translocation to the nucleus / Impaired BRCA2 binding to SEM1 (DSS1) / meiosis I / proteasome accessory complex / purine ribonucleoside triphosphate binding / integrator complex / proteasome regulatory particle / CD8-positive, alpha-beta T cell differentiation / thymic T cell selection / CD8-positive, alpha-beta T cell homeostasis / cytosolic proteasome complex / positive regulation of proteasomal protein catabolic process / proteasome-activating activity / Antigen processing: Ub, ATP-independent proteasomal degradation / proteasome regulatory particle, lid subcomplex / proteasome regulatory particle, base subcomplex / T-helper 1 cell differentiation / negative regulation of regulatory T cell differentiation / cellular response to type I interferon / protein K63-linked deubiquitination / metal-dependent deubiquitinase activity / negative regulation of programmed cell death / Regulation of ornithine decarboxylase (ODC) / proteasome core complex / Proteasome assembly / T-helper 17 cell differentiation / Cross-presentation of soluble exogenous antigens (endosomes) / transcription factor binding / K63-linked deubiquitinase activity / Somitogenesis / Homologous DNA Pairing and Strand Exchange / Defective homologous recombination repair (HRR) due to BRCA1 loss of function / Defective HDR through Homologous Recombination Repair (HRR) due to PALB2 loss of BRCA1 binding function / Defective HDR through Homologous Recombination Repair (HRR) due to PALB2 loss of BRCA2/RAD51/RAD51C binding function / Resolution of D-loop Structures through Synthesis-Dependent Strand Annealing (SDSA) / flagellated sperm motility / Resolution of D-loop Structures through Holliday Junction Intermediates / proteasome binding / Impaired BRCA2 binding to RAD51 / regulation of protein catabolic process / myofibril / AMPK-induced ERAD and lysosome mediated degradation of PD-L1(CD274) / GSK3B-mediated proteasomal degradation of PD-L1(CD274) / SPOP-mediated proteasomal degradation of PD-L1(CD274) / proteasomal ubiquitin-independent protein catabolic process / Ribosome Quality Control (RQC) complex extracts and degrades nascent peptide / general transcription initiation factor binding / proteasome storage granule / Presynaptic phase of homologous DNA pairing and strand exchange / protein deubiquitination / polyubiquitin modification-dependent protein binding / proteasome endopeptidase complex / NF-kappaB binding / proteasome core complex, beta-subunit complex / endopeptidase activator activity / threonine-type endopeptidase activity / mRNA export from nucleus / proteasome core complex, alpha-subunit complex / proteasome assembly / immune system process / regulation of G1/S transition of mitotic cell cycle / enzyme regulator activity / ciliary tip / response to type II interferon / positive regulation of interleukin-2 production / inclusion body / regulation of proteasomal protein catabolic process / TBP-class protein binding / : / proteasome complex / stem cell differentiation / sarcomere / proteasomal protein catabolic process / Regulation of activated PAK-2p34 by proteasome mediated degradation / sperm end piece / ubiquitin binding / Autodegradation of Cdh1 by Cdh1:APC/C / APC/C:Cdc20 mediated degradation of Securin / negative regulation of inflammatory response to antigenic stimulus / N-glycan trimming in the ER and Calnexin/Calreticulin cycle / Asymmetric localization of PCP proteins / Ubiquitin-dependent degradation of Cyclin D / lipopolysaccharide binding / SCF-beta-TrCP mediated degradation of Emi1 / NIK-->noncanonical NF-kB signaling / AUF1 (hnRNP D0) binds and destabilizes mRNA / TNFR2 non-canonical NF-kB pathway / Assembly of the pre-replicative complex / Vpu mediated degradation of CD4 / P-body / Cdc20:Phospho-APC/C mediated degradation of Cyclin A / Dectin-1 mediated noncanonical NF-kB signaling / Degradation of DVL / Degradation of AXIN / Degradation of CRY and PER proteins / meiotic cell cycle Similarity search - Function | |||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | |||||||||
| Method | single particle reconstruction / cryo EM / Resolution: 3.84 Å | |||||||||
 Authors Authors | Peddada N / Beutler B | |||||||||
| Funding support |  United States, 2 items United States, 2 items
| |||||||||
 Citation Citation | Journal: Proc Natl Acad Sci U S A / Year: 2025 Title: Structural insights into the ubiquitin-independent midnolin-proteasome pathway. Authors: Nagesh Peddada / Xue Zhong / Yan Yin / Danielle Renee Lazaro / Jianhui Wang / Stephen Lyon / Jin Huk Choi / Xiao-Chen Bai / Eva Marie Y Moresco / Bruce Beutler / Abstract: The protein midnolin (MIDN) augments proteasome activity in lymphocytes and dramatically facilitates the survival and proliferation of B-lymphoid malignancies. MIDN binds both to proteasomes and to ...The protein midnolin (MIDN) augments proteasome activity in lymphocytes and dramatically facilitates the survival and proliferation of B-lymphoid malignancies. MIDN binds both to proteasomes and to substrates, but the mode of interaction with the proteasome is unknown, and the mechanism by which MIDN facilitates substrate degradation in a ubiquitin-independent manner is incompletely understood. Here, we present cryoelectron microscopy (cryo-EM) structures of the substrate-engaged, MIDN-bound human proteasome in two conformational states. MIDN induces proteasome conformations similarly to ubiquitinated substrates by using its ubiquitin-like domain to bind to the deubiquitinase RPN11 (PSMD14). By simultaneously binding to RPN1 (PSMD2) with its C-terminal α-helix, MIDN positions its substrate-carrying Catch domain above the proteasome ATPase channel through which substrates are translocated before degradation. Our findings suggest that both ubiquitin-like domain and C-terminal α-helix must bind to the proteasome for MIDN to stimulate proteasome activity. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Supplemental images |
|---|

- Downloads & links
Downloads & links
-EMDB archive
| Map data | emd_49510.map.gz | 473.8 MB | EMDB map data format | |
|---|---|---|---|---|
| Header (meta data) | emd-49510-v30.xmlemd-49510.xml | 57.1 KB 57.1 KB | Display Display | EMDB header |
| Images |  emd_49510.png emd_49510.png | 81.4 KB | ||
| Filedesc metadata | emd-49510.cif.gz | 15.2 KB | ||
| Archive directory |  http://ftp.pdbj.org/pub/emdb/structures/EMD-49510ftp://ftp.pdbj.org/pub/emdb/structures/EMD-49510 http://ftp.pdbj.org/pub/emdb/structures/EMD-49510ftp://ftp.pdbj.org/pub/emdb/structures/EMD-49510 | HTTPS FTP |
-Related structure data
| Related structure data | 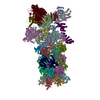 9nkjMC 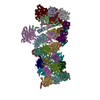 9nkfC  9nkgC  9nkiC C: citing same article ( M: atomic model generated by this map |
|---|---|
| Similar structure data |













-Links
| EMDB pages | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| Related items in Molecule of the Month |












-Map
| File | Download / File: emd_49510.map.gz / Format: CCP4 / Size: 1000 MB / Type: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Projections & slices | Image control
Images are generated by Spider. | ||||||||||||||||||||||||||||||||||||
| Voxel size | X=Y=Z: 1.074 Å | ||||||||||||||||||||||||||||||||||||
| Density |
| ||||||||||||||||||||||||||||||||||||
| Symmetry | Space group: 1 | ||||||||||||||||||||||||||||||||||||
| Details | EMDB XML:
|
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-Supplemental data
- Sample components
Sample components
+Entire : In Vitro Reconstituted MIDN-26S Proteasome complex
+Supramolecule #1: In Vitro Reconstituted MIDN-26S Proteasome complex
+Macromolecule #1: 26S proteasome regulatory subunit 7
+Macromolecule #2: 26S proteasome regulatory subunit 4
+Macromolecule #3: 26S protease regulatory subunit 8
+Macromolecule #4: 26S proteasome regulatory subunit 6B
+Macromolecule #5: 26S proteasome regulatory subunit 10B
+Macromolecule #6: 26S proteasome regulatory subunit 6A
+Macromolecule #7: 26S proteasome non-ATPase regulatory subunit 1
+Macromolecule #8: 26S proteasome non-ATPase regulatory subunit 3
+Macromolecule #9: 26S proteasome non-ATPase regulatory subunit 11
+Macromolecule #10: 26S proteasome non-ATPase regulatory subunit 6
+Macromolecule #11: 26S proteasome non-ATPase regulatory subunit 7
+Macromolecule #12: 26S proteasome non-ATPase regulatory subunit 13
+Macromolecule #13: 26S proteasome non-ATPase regulatory subunit 4
+Macromolecule #14: 26S proteasome non-ATPase regulatory subunit 14
+Macromolecule #15: 26S proteasome non-ATPase regulatory subunit 8
+Macromolecule #16: 26S proteasome non-ATPase regulatory subunit 2
+Macromolecule #17: unknown density-Substrate density
+Macromolecule #18: 26S proteasome non-ATPase regulatory subunit 12
+Macromolecule #19: 26S proteasome complex subunit SEM1
+Macromolecule #20: Proteasome subunit alpha type-6
+Macromolecule #21: Proteasome subunit alpha type-2
+Macromolecule #22: Proteasome subunit alpha type-4
+Macromolecule #23: Proteasome subunit alpha type-7
+Macromolecule #24: Proteasome subunit alpha type-5
+Macromolecule #25: Proteasome subunit alpha type-1
+Macromolecule #26: Proteasome subunit alpha type-3
+Macromolecule #27: Proteasome subunit beta type-6
+Macromolecule #28: Proteasome subunit beta type-7
+Macromolecule #29: Proteasome subunit beta type-3
+Macromolecule #30: Proteasome subunit beta type-2
+Macromolecule #31: Proteasome subunit beta type-5
+Macromolecule #32: Proteasome subunit beta type-1
+Macromolecule #33: Proteasome subunit beta type-4
+Macromolecule #34: ADENOSINE-5'-TRIPHOSPHATE

+Macromolecule #35: MAGNESIUM ION
+Macromolecule #36: ADENOSINE-5'-DIPHOSPHATE

+Macromolecule #37: ZINC ION
+Macromolecule #38: N-[(benzyloxy)carbonyl]-L-leucyl-N-[(2S)-4-methyl-1-oxopentan-2-y...

-Experimental details
-Structure determination
| Method | cryo EM |
|---|---|
 Processing Processing | single particle reconstruction |
| Aggregation state | particle |
-Sample preparation
| Buffer | pH: 7.6 Details: 50 mM Tris pH 7.5, 150 mM NaCl, 20 mM KCl, 5 mM MgCl2, and 1 mM TECP |
|---|---|
| Grid | Model: Quantifoil R1.2/1.3 / Material: COPPER / Mesh: 300 / Pretreatment - Type: GLOW DISCHARGE |
| Vitrification | Cryogen name: ETHANE / Instrument: FEI VITROBOT MARK IV |
- Electron microscopy
Electron microscopy
| Microscope | TFS KRIOS |
|---|---|
| Software | Name: SerialEM |
| Image recording | Film or detector model: GATAN K3 BIOQUANTUM (6k x 4k) / Number grids imaged: 3 / Average electron dose: 50.0 e/Å2 |
| Electron beam | Acceleration voltage: 300 kV / Electron source:  FIELD EMISSION GUN FIELD EMISSION GUN |
| Electron optics | Illumination mode: FLOOD BEAM / Imaging mode: BRIGHT FIELD / Nominal defocus max: 2.7 µm / Nominal defocus min: 1.2 µm |
| Sample stage | Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |