 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 |  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
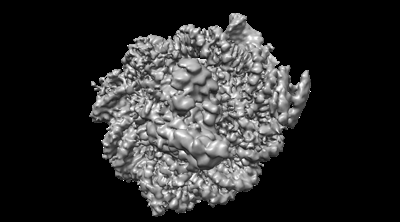
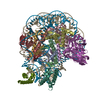
| タイトル | Structure of LIN28b nucleosome bound 2 OCT4 | |||||||||
 マップデータ マップデータ | ||||||||||
 試料 試料 |
| |||||||||
 キーワード キーワード | nucleosome / transcription factor / transcription / CHROMATIN BINDING PROTEIN-DNA complex / TRANSCRIPTION-DNA complex | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報cell fate commitment involved in formation of primary germ layer / cardiac cell fate determination / POU5F1 (OCT4), SOX2, NANOG repress genes related to differentiation / Formation of the anterior neural plate / endodermal-mesodermal cell signaling / regulation of asymmetric cell division / endodermal cell fate specification / heart induction / POU5F1 (OCT4), SOX2, NANOG activate genes related to proliferation / Specification of the neural plate border ...cell fate commitment involved in formation of primary germ layer / cardiac cell fate determination / POU5F1 (OCT4), SOX2, NANOG repress genes related to differentiation / Formation of the anterior neural plate / endodermal-mesodermal cell signaling / regulation of asymmetric cell division / endodermal cell fate specification / heart induction / POU5F1 (OCT4), SOX2, NANOG activate genes related to proliferation / Specification of the neural plate border / Specification of primordial germ cells / Transcriptional regulation of pluripotent stem cells / Germ layer formation at gastrulation / miRNA binding / somatic stem cell population maintenance / carbohydrate transmembrane transporter activity / anatomical structure morphogenesis / blastocyst development / BMP signaling pathway / negative regulation of megakaryocyte differentiation / protein localization to CENP-A containing chromatin / Chromatin modifying enzymes / Replacement of protamines by nucleosomes in the male pronucleus / CENP-A containing nucleosome / Packaging Of Telomere Ends / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / Deposition of new CENPA-containing nucleosomes at the centromere / epigenetic regulation of gene expression / telomere organization / negative regulation of miRNA transcription / Interleukin-7 signaling / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / RNA Polymerase I Promoter Opening / Inhibition of DNA recombination at telomere / Assembly of the ORC complex at the origin of replication / Meiotic synapsis / SUMOylation of chromatin organization proteins / Regulation of endogenous retroelements by the Human Silencing Hub (HUSH) complex / DNA methylation / Condensation of Prophase Chromosomes / Chromatin modifications during the maternal to zygotic transition (MZT) / SIRT1 negatively regulates rRNA expression / HCMV Late Events / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / PRC2 methylates histones and DNA / innate immune response in mucosa / Regulation of endogenous retroelements by KRAB-ZFP proteins / Defective pyroptosis / HDACs deacetylate histones / Regulation of endogenous retroelements by Piwi-interacting RNAs (piRNAs) / Nonhomologous End-Joining (NHEJ) / RNA Polymerase I Promoter Escape / Transcriptional regulation by small RNAs / Formation of the beta-catenin:TCF transactivating complex / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / HDMs demethylate histones / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / G2/M DNA damage checkpoint / Negative Regulation of CDH1 Gene Transcription / NoRC negatively regulates rRNA expression / PKMTs methylate histone lysines / B-WICH complex positively regulates rRNA expression / response to wounding / DNA Damage/Telomere Stress Induced Senescence / DNA-binding transcription repressor activity, RNA polymerase II-specific / Pre-NOTCH Transcription and Translation / Meiotic recombination / Activation of anterior HOX genes in hindbrain development during early embryogenesis / Transcriptional regulation of granulopoiesis / Metalloprotease DUBs / RMTs methylate histone arginines / HCMV Early Events / sequence-specific double-stranded DNA binding / structural constituent of chromatin / UCH proteinases / positive regulation of canonical Wnt signaling pathway / nucleosome / antimicrobial humoral immune response mediated by antimicrobial peptide / heterochromatin formation / nucleosome assembly / antibacterial humoral response / HATs acetylate histones / outer membrane-bounded periplasmic space / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / Factors involved in megakaryocyte development and platelet production / MLL4 and MLL3 complexes regulate expression of PPARG target genes in adipogenesis and hepatic steatosis / chromatin organization / RUNX1 regulates transcription of genes involved in differentiation of HSCs / Processing of DNA double-strand break ends / regulation of gene expression / Senescence-Associated Secretory Phenotype (SASP) / Oxidative Stress Induced Senescence / transcription regulator complex / Estrogen-dependent gene expression / sequence-specific DNA binding / RNA polymerase II-specific DNA-binding transcription factor binding / DNA-binding transcription factor activity, RNA polymerase II-specific / chromosome, telomeric region 類似検索 - 分子機能 | |||||||||
| 生物種 |  Homo sapiens (ヒト) / Homo sapiens (ヒト) /  | |||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 2.6 Å | |||||||||
 データ登録者 データ登録者 | Lian T / Guan R / Bai Y | |||||||||
| 資金援助 |  米国, 1件 米国, 1件
| |||||||||
 引用 引用 | ジャーナル: Mol Cell / 年: 2023 タイトル: Structural mechanism of LIN28B nucleosome targeting by OCT4. 著者: Ruifang Guan / Tengfei Lian / Bing-Rui Zhou / David Wheeler / Yawen Bai / 要旨: Pioneer transcription factors are essential for cell fate changes by targeting closed chromatin. OCT4 is a crucial pioneer factor that can induce cell reprogramming. However, the structural basis of ...Pioneer transcription factors are essential for cell fate changes by targeting closed chromatin. OCT4 is a crucial pioneer factor that can induce cell reprogramming. However, the structural basis of how pioneer factors recognize the in vivo nucleosomal DNA targets is unknown. Here, we determine the high-resolution structures of the nucleosome containing human LIN28B DNA and its complexes with the OCT4 DNA binding region. Three OCT4s bind the pre-positioned nucleosome by recognizing non-canonical DNA sequences. Two use their POUS domains while the other uses the POUS-loop-POUHD region; POUHD serves as a wedge to unwrap ∼25 base pair DNA. Our analysis of previous genomic data and determination of the ESRRB-nucleosome-OCT4 structure confirmed the generality of these structural features. Moreover, biochemical studies suggest that multiple OCT4s cooperatively open the H1-condensed nucleosome array containing the LIN28B nucleosome. Thus, our study suggests a mechanism of how OCT4 can target the nucleosome and open closed chromatin. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 添付画像 |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_26260.map.gz | 34.2 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-26260-v30.xmlemd-26260.xml | 18 KB 18 KB | 表示 表示 | EMDBヘッダ |
| 画像 |  emd_26260.png emd_26260.png | 54.4 KB | ||
| Filedesc metadata | emd-26260.cif.gz | 6.9 KB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-26260ftp://ftp.pdbj.org/pub/emdb/structures/EMD-26260 http://ftp.pdbj.org/pub/emdb/structures/EMD-26260ftp://ftp.pdbj.org/pub/emdb/structures/EMD-26260 | HTTPS FTP |
-関連構造データ





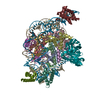
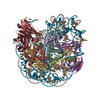
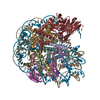
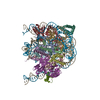
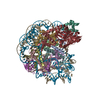
-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| 「今月の分子」の関連する項目 |













-マップ
| ファイル | ダウンロード / ファイル: emd_26260.map.gz / 形式: CCP4 / 大きさ: 52.7 MB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 投影像・断面図 | 画像のコントロール
画像は Spider により作成 | ||||||||||||||||||||||||||||||||||||
| ボクセルのサイズ | X=Y=Z: 1.056 Å | ||||||||||||||||||||||||||||||||||||
| 密度 |
| ||||||||||||||||||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||||||||||||||||||
| 詳細 | EMDB XML:
|
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-添付データ
- 試料の構成要素
試料の構成要素
-全体 : Complex of nucleosome bound to two OCT-4s
| 全体 | 名称: Complex of nucleosome bound to two OCT-4s |
|---|---|
| 要素 |
|
-超分子 #1: Complex of nucleosome bound to two OCT-4s
| 超分子 | 名称: Complex of nucleosome bound to two OCT-4s / タイプ: complex / ID: 1 / 親要素: 0 / 含まれる分子: #1-#8 |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
-分子 #1: Histone H3.1
| 分子 | 名称: Histone H3.1 / タイプ: protein_or_peptide / ID: 1 / コピー数: 2 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 15.437167 KDa |
| 組換発現 | 生物種:  |
| 配列 | 文字列: MARTKQTARK STGGKAPRKQ LATKAARKSA PATGGVKKPH RYRPGTVALR EIRRYQKSTE LLIRKLPFQR LVREIAQDFK TDLRFQSSA VMALQEACEA YLVGLFEDTN LCAIHAKRVT IMPKDIQLAR RIRGERA UniProtKB: Histone H3.1 |
-分子 #2: Histone H4
| 分子 | 名称: Histone H4 / タイプ: protein_or_peptide / ID: 2 / コピー数: 2 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 11.394426 KDa |
| 組換発現 | 生物種: |
| 配列 | 文字列: MSGRGKGGKG LGKGGAKRHR KVLRDNIQGI TKPAIRRLAR RGGVKRISGL IYEETRGVLK VFLENVIRDA VTYTEHAKRK TVTAMDVVY ALKRQGRTLY GFGG UniProtKB: Histone H4 |
-分子 #3: Histone H2A type 2-C
| 分子 | 名称: Histone H2A type 2-C / タイプ: protein_or_peptide / ID: 3 / コピー数: 2 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 14.017428 KDa |
| 組換発現 | 生物種: |
| 配列 | 文字列: MSGRGKQGGK ARAKAKSRSS RAGLQFPVGR VHRLLRKGNY AERVGAGAPV YMAAVLEYLT AEILELAGNA ARDNKKTRII PRHLQLAIR NDEELNKLLG KVTIAQGGVL PNIQAVLLPK KTESHKAKSK UniProtKB: Histone H2A type 2-C |
-分子 #4: Histone H2B type 2-E
| 分子 | 名称: Histone H2B type 2-E / タイプ: protein_or_peptide / ID: 4 / コピー数: 2 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 13.951239 KDa |
| 組換発現 | 生物種: |
| 配列 | 文字列: MPEPAKSAPA PKKGSKKAVT KAQKKDGKKR KRSRKESYSI YVYKVLKQVH PDTGISSKAM GIMNSFVNDI FERIAGEASR LAHYNKRST ITSREIQTAV RLLLPGELAK HAVSEGTKAV TKYTSSK UniProtKB: Histone H2B type 2-E |
-分子 #7: Maltodextrin-binding protein,POU domain, class 5, transcription f...
| 分子 | 名称: Maltodextrin-binding protein,POU domain, class 5, transcription factor 1 タイプ: protein_or_peptide / ID: 7 / コピー数: 2 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 61.269703 KDa |
| 組換発現 | 生物種: |
| 配列 | 文字列: MKEAKIEEGK LVIWINGDKG YNGLAEVGKK FEKDTGIKVT VEHPDKLEEK FPQVAATGDG PDIIFWAHDR FGGYAQSGLL AEITPDKAF QDKLYPFTWD AVRYNGKLIA YPIAVEALSL IYNKDLLPNP PKTWEEIPAL DKELKAKGKS ALMFNLQEPY F TWPLIAAD ...文字列: MKEAKIEEGK LVIWINGDKG YNGLAEVGKK FEKDTGIKVT VEHPDKLEEK FPQVAATGDG PDIIFWAHDR FGGYAQSGLL AEITPDKAF QDKLYPFTWD AVRYNGKLIA YPIAVEALSL IYNKDLLPNP PKTWEEIPAL DKELKAKGKS ALMFNLQEPY F TWPLIAAD GGYAFKYENG KYDIKDVGVD NAGAKAGLTF LVDLIKNKHM NADTDYSIAE AAFNKGETAM TINGPWAWSN ID TSKVNYG VTVLP(UNK)FKGQ PSKPFVGVLS AGINAASPNK ELAKEFLENY LLTDEGLEAV NKDKPLGAVA LKSYEEELA KDPRIAATME NAQKGEIMPN IPQMSAFWYA VRTAVINAAS GRQTVDAALA AAQTNAGSEN LYFQGSVDSA AASDIKALQK ELEQFAKLL KQKRITLGYT QADVGLTLGV LFGKVFSQTT ICRFEALQLS FKNMCKLRPL LQKWVEEADN NENLQEICKA E TLVQARKR KRTSIENRVR GNLENLFLQC PKPTLQQISH IAQQLGLEKD VVRVWFCNRR QKGKRSSSEF HHHHHH UniProtKB: Maltodextrin-binding protein, POU domain, class 5, transcription factor 1 |
-分子 #8: Single-chain variable fragment
| 分子 | 名称: Single-chain variable fragment / タイプ: protein_or_peptide / ID: 8 / コピー数: 2 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: |
| 分子量 | 理論値: 29.030146 KDa |
| 組換発現 | 生物種: |
| 配列 | 文字列: MKSSHHHHHH ENLYFQSNAM EVQLQQSGPE LVEPGTSVKM PCKASGYTFT SYTIQWVKQT PRQGLEWIGY IYPYNAGTKY NEKFKGKAT LTSDKSSSTV YMELSSLTSE DSAVYYCARK SSRLRSTLDY WGQGTSVTVS SGGGGSGGGG SGGGGSMDIK M TQSPSSMH ...文字列: MKSSHHHHHH ENLYFQSNAM EVQLQQSGPE LVEPGTSVKM PCKASGYTFT SYTIQWVKQT PRQGLEWIGY IYPYNAGTKY NEKFKGKAT LTSDKSSSTV YMELSSLTSE DSAVYYCARK SSRLRSTLDY WGQGTSVTVS SGGGGSGGGG SGGGGSMDIK M TQSPSSMH ASLGERVTIT CKASQDIRSY LSWYQQKPWK SPKTLIYYAT SLADGVPSRF SGSGSGQDFS LTINNLESDD TA TYYCLQH GESPYTFGSG TKLEIKRA |
-分子 #5: DNA (162-MER)
| 分子 | 名称: DNA (162-MER) / タイプ: dna / ID: 5 / コピー数: 1 / 分類: DNA |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 50.398344 KDa |
| 配列 | 文字列: (DA)(DG)(DT)(DG)(DG)(DT)(DA)(DT)(DT)(DA) (DA)(DC)(DA)(DT)(DA)(DT)(DC)(DC)(DT)(DC) (DA)(DG)(DT)(DG)(DG)(DT)(DG)(DA)(DG) (DT)(DA)(DT)(DT)(DA)(DA)(DC)(DA)(DT)(DG) (DG) (DA)(DA)(DC)(DT)(DT) ...文字列: (DA)(DG)(DT)(DG)(DG)(DT)(DA)(DT)(DT)(DA) (DA)(DC)(DA)(DT)(DA)(DT)(DC)(DC)(DT)(DC) (DA)(DG)(DT)(DG)(DG)(DT)(DG)(DA)(DG) (DT)(DA)(DT)(DT)(DA)(DA)(DC)(DA)(DT)(DG) (DG) (DA)(DA)(DC)(DT)(DT)(DA)(DC)(DT) (DC)(DC)(DA)(DA)(DC)(DA)(DA)(DT)(DA)(DC) (DA)(DG) (DA)(DT)(DG)(DC)(DT)(DG)(DA) (DA)(DT)(DA)(DA)(DA)(DT)(DG)(DT)(DA)(DG) (DT)(DC)(DT) (DA)(DA)(DG)(DT)(DG)(DA) (DA)(DG)(DG)(DA)(DA)(DG)(DA)(DA)(DG)(DG) (DA)(DA)(DA)(DG) (DG)(DT)(DG)(DG)(DG) (DA)(DG)(DC)(DT)(DG)(DC)(DC)(DA)(DT)(DC) (DA)(DC)(DT)(DC)(DA) (DG)(DA)(DA)(DT) (DT)(DG)(DT)(DC)(DC)(DA)(DG)(DC)(DA)(DG) (DG)(DG)(DA)(DT)(DT)(DG) (DT)(DG)(DC) (DA)(DA)(DG)(DC)(DT)(DT)(DG)(DT)(DG)(DA) (DA)(DT)(DA)(DA)(DA)(DG)(DA) (DC)(DA) GENBANK: GENBANK: Z95329.1 |
-分子 #6: DNA (162-MER)
| 分子 | 名称: DNA (162-MER) / タイプ: dna / ID: 6 / コピー数: 1 / 分類: DNA |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 49.59568 KDa |
| 配列 | 文字列: (DT)(DG)(DT)(DC)(DT)(DT)(DT)(DA)(DT)(DT) (DC)(DA)(DC)(DA)(DA)(DG)(DC)(DT)(DT)(DG) (DC)(DA)(DC)(DA)(DA)(DT)(DC)(DC)(DC) (DT)(DG)(DC)(DT)(DG)(DG)(DA)(DC)(DA)(DA) (DT) (DT)(DC)(DT)(DG)(DA) ...文字列: (DT)(DG)(DT)(DC)(DT)(DT)(DT)(DA)(DT)(DT) (DC)(DA)(DC)(DA)(DA)(DG)(DC)(DT)(DT)(DG) (DC)(DA)(DC)(DA)(DA)(DT)(DC)(DC)(DC) (DT)(DG)(DC)(DT)(DG)(DG)(DA)(DC)(DA)(DA) (DT) (DT)(DC)(DT)(DG)(DA)(DG)(DT)(DG) (DA)(DT)(DG)(DG)(DC)(DA)(DG)(DC)(DT)(DC) (DC)(DC) (DA)(DC)(DC)(DT)(DT)(DT)(DC) (DC)(DT)(DT)(DC)(DT)(DT)(DC)(DC)(DT)(DT) (DC)(DA)(DC) (DT)(DT)(DA)(DG)(DA)(DC) (DT)(DA)(DC)(DA)(DT)(DT)(DT)(DA)(DT)(DT) (DC)(DA)(DG)(DC) (DA)(DT)(DC)(DT)(DG) (DT)(DA)(DT)(DT)(DG)(DT)(DT)(DG)(DG)(DA) (DG)(DT)(DA)(DA)(DG) (DT)(DT)(DC)(DC) (DA)(DT)(DG)(DT)(DT)(DA)(DA)(DT)(DA)(DC) (DT)(DC)(DA)(DC)(DC)(DA) (DC)(DT)(DG) (DA)(DG)(DG)(DA)(DT)(DA)(DT)(DG)(DT)(DT) (DA)(DA)(DT)(DA)(DC)(DC)(DA) (DC)(DT) GENBANK: GENBANK: Z95329.1 |
-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | particle |
-試料調製
| 緩衝液 | pH: 7.3 |
|---|---|
| 凍結 | 凍結剤: ETHANE |
- 電子顕微鏡法
電子顕微鏡法
| 顕微鏡 | FEI TITAN KRIOS |
|---|---|
| 撮影 | フィルム・検出器のモデル: GATAN K3 (6k x 4k) / 平均電子線量: 53.8 e/Å2 |
| 電子線 | 加速電圧: 300 kV / 電子線源:  FIELD EMISSION GUN FIELD EMISSION GUN |
| 電子光学系 | 照射モード: SPOT SCAN / 撮影モード: BRIGHT FIELD / 最大 デフォーカス(公称値): 2.0 µm / 最小 デフォーカス(公称値): 1.0 µm |
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
-画像解析
| 初期モデル | モデルのタイプ: NONE |
|---|---|
| 最終 再構成 | 解像度のタイプ: BY AUTHOR / 解像度: 2.6 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 使用した粒子像数: 88819 |
| 初期 角度割当 | タイプ: MAXIMUM LIKELIHOOD |
| 最終 角度割当 | タイプ: MAXIMUM LIKELIHOOD |
