 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: EMDB / ID: EMD-23829 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
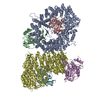
| タイトル | Structure of yeast cytoplasmic dynein with AAA3 Walker B mutation bound to Lis1 | |||||||||
 マップデータ マップデータ | scDynein E2488Q bound to two Lis1s | |||||||||
 試料 試料 |
| |||||||||
 キーワード キーワード | motor / AAA / MOTOR PROTEIN | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報microtubule sliding / microtubule organizing center organization / nuclear migration along microtubule / microtubule plus-end binding / vesicle transport along microtubule / dynein complex / minus-end-directed microtubule motor activity / microtubule associated complex / microtubule-based movement / nuclear migration ...microtubule sliding / microtubule organizing center organization / nuclear migration along microtubule / microtubule plus-end binding / vesicle transport along microtubule / dynein complex / minus-end-directed microtubule motor activity / microtubule associated complex / microtubule-based movement / nuclear migration / dynein complex binding / establishment of mitotic spindle orientation / Antigen processing: Ubiquitination & Proteasome degradation / cytoplasmic microtubule / kinetochore / spindle pole / nuclear envelope / microtubule / cell division / ATP binding / identical protein binding / nucleus / cytoplasm 類似検索 - 分子機能 | |||||||||
| 生物種 |  | |||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.1 Å | |||||||||
 データ登録者 データ登録者 | Lahiri I / Reimer JM | |||||||||
| 資金援助 |  米国, 1件 米国, 1件
| |||||||||
 引用 引用 | ジャーナル: Elife / 年: 2022 タイトル: Structural basis for cytoplasmic dynein-1 regulation by Lis1. 著者: John P Gillies / Janice M Reimer / Eva P Karasmanis / Indrajit Lahiri / Zaw Min Htet / Andres E Leschziner / Samara L Reck-Peterson /  要旨: The lissencephaly 1 gene, , is mutated in patients with the neurodevelopmental disease lissencephaly. The Lis1 protein is conserved from fungi to mammals and is a key regulator of cytoplasmic dynein- ...The lissencephaly 1 gene, , is mutated in patients with the neurodevelopmental disease lissencephaly. The Lis1 protein is conserved from fungi to mammals and is a key regulator of cytoplasmic dynein-1, the major minus-end-directed microtubule motor in many eukaryotes. Lis1 is the only dynein regulator known to bind directly to dynein's motor domain, and by doing so alters dynein's mechanochemistry. Lis1 is required for the formation of fully active dynein complexes, which also contain essential cofactors: dynactin and an activating adaptor. Here, we report the first high-resolution structure of the yeast dynein-Lis1 complex. Our 3.1 Å structure reveals, in molecular detail, the major contacts between dynein and Lis1 and between Lis1's ß-propellers. Structure-guided mutations in Lis1 and dynein show that these contacts are required for Lis1's ability to form fully active human dynein complexes and to regulate yeast dynein's mechanochemistry and in vivo function. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | EMマップ: SurfViewMolmilJmol/JSmol |
| 添付画像 |

- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_23829.map.gz | 93.9 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-23829-v30.xmlemd-23829.xml | 19.7 KB 19.7 KB | 表示 表示 | EMDBヘッダ |
| FSC (解像度算出) | emd_23829_fsc.xml | 10.6 KB | 表示 | FSCデータファイル |
| 画像 | 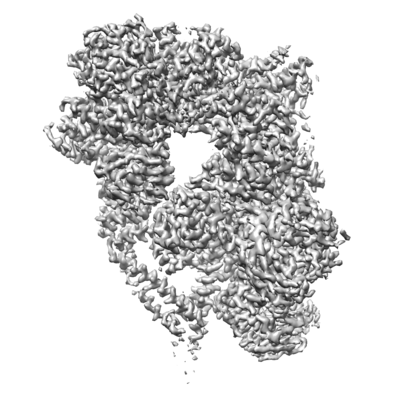 emd_23829.png emd_23829.png | 123.6 KB | ||
| Filedesc metadata | emd-23829.cif.gz | 7.7 KB | ||
| その他 | emd_23829_half_map_1.map.gzemd_23829_half_map_2.map.gz | 80.4 MB 80.4 MB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-23829ftp://ftp.pdbj.org/pub/emdb/structures/EMD-23829 http://ftp.pdbj.org/pub/emdb/structures/EMD-23829ftp://ftp.pdbj.org/pub/emdb/structures/EMD-23829 | HTTPS FTP |
-検証レポート
| 文書・要旨 | emd_23829_validation.pdf.gz | 940 KB | 表示 | EMDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | emd_23829_full_validation.pdf.gz | 939.5 KB | 表示 | |
| XML形式データ | emd_23829_validation.xml.gz | 17.8 KB | 表示 | |
| CIF形式データ | emd_23829_validation.cif.gz | 23.5 KB | 表示 | |
| アーカイブディレクトリ | https://ftp.pdbj.org/pub/emdb/validation_reports/EMD-23829ftp://ftp.pdbj.org/pub/emdb/validation_reports/EMD-23829 | HTTPS FTP |
-関連構造データ


-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| 「今月の分子」の関連する項目 |





-マップ
| ファイル | ダウンロード / ファイル: emd_23829.map.gz / 形式: CCP4 / 大きさ: 103 MB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | scDynein E2488Q bound to two Lis1s | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 投影像・断面図 | 画像のコントロール
画像は Spider により作成 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ボクセルのサイズ | X=Y=Z: 1.31 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 密度 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 詳細 | EMDB XML:
CCP4マップ ヘッダ情報:
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-添付データ
-ハーフマップ: Half map 1
| ファイル | emd_23829_half_map_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | Half map 1 | ||||||||||||
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |








-ハーフマップ: Half map 2
| ファイル | emd_23829_half_map_2.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | Half map 2 | ||||||||||||
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |








- 試料の構成要素
試料の構成要素
-全体 : Complex of yeast dynein bound by two Lis1s in the presence of ATP-Va
| 全体 | 名称: Complex of yeast dynein bound by two Lis1s in the presence of ATP-Va |
|---|---|
| 要素 |
|
-超分子 #1: Complex of yeast dynein bound by two Lis1s in the presence of ATP-Va
| 超分子 | 名称: Complex of yeast dynein bound by two Lis1s in the presence of ATP-Va タイプ: complex / ID: 1 / 親要素: 0 / 含まれる分子: #1-#2 |
|---|---|
| 由来(天然) | 生物種: |
-分子 #1: dynein AAA3-WalkerB mutant (E2488Q)
| 分子 | 名称: dynein AAA3-WalkerB mutant (E2488Q) / タイプ: protein_or_peptide / ID: 1 / コピー数: 1 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: |
| 分子量 | 理論値: 331.524 KDa |
| 配列 | 文字列: GDQLTHVVEE VKTYDLVWRS IKNLWEDVQR TFETPWCRVD VLLLQSDLAN FLRRADELPR AVKQFEMYKS LFSQVNMLTS VNKILVELK DGALKPRHWN MIFRDIGKRQ IQKNLLDKLE FSLKDVMVLN LTLNEILLTK IIERAQKEFV IEKSLNRIKK F WKEAQYEV ...文字列: GDQLTHVVEE VKTYDLVWRS IKNLWEDVQR TFETPWCRVD VLLLQSDLAN FLRRADELPR AVKQFEMYKS LFSQVNMLTS VNKILVELK DGALKPRHWN MIFRDIGKRQ IQKNLLDKLE FSLKDVMVLN LTLNEILLTK IIERAQKEFV IEKSLNRIKK F WKEAQYEV IEHSSGLKLV REWDVLEQAC KEDLEELVSM KASNYYKIFE QDCLDLESKL TKLSEIQVNW VEVQFYWLDL YG ILGENLD IQNFLPLETS KFKSLTSEYK MITTRAFQLD TTIEVIHIPN FDTTLKLTID SLKMIKSSLS TFLERQRRQF PRF YFLGND DLLKIIGSGK HHDQVSKFMK KMFGSIESII FFEDSITGVR SVEGEVLNLN EKIELKDSIQ AQEWLNILDT EIKL SVFTQ FRDCLGQLKD GTDIEVVVSK YIFQAILLSA QVMWTELVEK CLQTNEFSKY WKEVDMKIKG LLDKLNKSSD NVKKK IEAL LVEYLHFNNV IGQLKNCSTK EEARLLWAKV QKFYQKNDTL DDLNSVFISQ SGYLLQYKFE YIGIPERLIY TPLLLV GFA TLTDSLHQKY GGCFFGPAGT GKTETVKAFG QNLGRVVVVF NCDDSFDYQV LSRLLVGITQ IGAWGCFDEF NRLDEKV LS AVSANIQQIQ NGLQVGKSHI TLLEEETPLS PHTAVFITLN PGYNGRSELP ENLKKSFREF SMKSPQSGTI AEMILQIM G FEDSKSLASK IVHFLELLSS KCSSMNHYHF GLRTLKGVLR NCSPLVSEFG EGEKTVVESL KRVILPSLGD TDELVFKDE LSKIFDSAGT PLNSKAIVQC LKDAGQRSGF SMSEEFLKKC MQFYYMQKTQ QALILVGKAG CGKTATWKTV IDAMAIFDGH ANVVYVIDT KVLTKESLYG SMLKATLEWR DGLFTSILRR VNDDITGTFK NSRIWVVFDS DLDPEYVEAM NSVLDDNKIL T LPNGERLP IPPNFRILFE TDNLDHTTPA TITRCGLLWF STDVCSISSK IDHLLNKSYE ALDNKLSMFE LDKLKDLISD SF DMASLTN IFTCSNDLVH ILGVRTFNKL ETAVQLAVHL ISSYRQWFQN LDDKSLKDVI TLLIKRSLLY ALAGDSTGES QRA FIQTIN TYFGHDSQEL SDYSTIVIAN DKLSFSSFCS EIPSVSLEAH EVMRPDIVIP TIDTIKHEKI FYDLLNSKRG IILC GPPGS GKTMIMNNAL RNSSLYDVVG INFSKDTTTE HILSALHRHT NYVTTSKGLT LLPKSDIKNL VLFCDQINLP KLDKY GSQN VVLFLRQLME KQGFWKTPEN KWVTIERIHI VGACNPPTDP GRIPMSERFT RHAAILYLGY PSGKSLSQIY EIYYKA IFK LVPEFRSYTE PFARASVHLY NECKARYSTG LQSHYLFSPR ELTRLVRGVY TAINTGPRQT LRSLIRLWAY EAWRIFA DR LVGVKEKNSF EQLLYETVDK YLPNQDLGNI SSTSLLFSGL LSLDFKEVNK TDLVNFIEER FKTFCDEELE VPMVIHES M VDHILRIDRA LKQVQGHMML IGASRTGKTI LTRFVAWLNG LKIVQPKIHR HSNLSDFDMI LKKAISDCSL KESRTCLII DESNILETAF LERMNTLLAN ADIPDLFQGE EYDKLLNNLR NKTRSLGLLL DTEQELYDWF VGEIAKNLHV VFTICDPTNN KSSAMISSP ALFNRCIINW MGDWDTKTMS QVANNMVDVV PMEFTDFIVP EVNKELVFTE PIQTIRDAVV NILIHFDRNF Y QKMKVGVN PRSPGYFIDG LRALVKLVTA KYQDLQENQR FVNVGLEKLN ESVLKVNELN KTLSKKSTEL TEKEKEARST LD KMLMEQN ESERKQEATE EIKKILKVQE EDIRKRKEVV MKSIQDIEPT ILEAQRGVKN IKKQQLTEIR SMVNPPSGVK IVM EAVCAI LGYQFSNWRD IQQFIRKDDF IHNIVHYDTT LHMKPQIRKY MEEEFLSDPN FTYETINRAS KACGPLYQWV NAQI NFSKV LENVDPLRQE MKRIEFESLK TKANLLAAEE MTQDLEASIE VSKQKYSLLI RDVEAIKTEM SNVQANLDRS ISLVK SLTF EKERWLNTTK QFSKTSQELI GNCIISSIYE TYFGHLNERE RGDMLVILKR LLGKFAVKYD VNYRFIDYLV TLDEKM KWL ECGLDKNDYF LENMSIVMNS QDAVPFLLDP SSHMITVISN YYGNKTVLLS FLEEGFVKRL ENAVRFGSVV IIQDGEF FD PIISRLISRE FNHAGNRVTV EIGDHEVDVS GDFKLFIHSC DPSGDIPIFL RSRVRLVHFV TNKESIETRI FDITLTEE N AEMQRKREDL IKLNTEYRLK LKNLEKRLLE ELNNSQGNML ENDELMVTLN NLKKEAMNIE KKLSESEEFF PQFDNLVEE YSIIGKHSVK IFSMLEKFGQ FHWFYGISIG QFLSCFKRVF IKKSRETRAA RTRVDEILWL LYQEVYCQFS TALDKKFKMI MAMTMFCLY KFDIESEQYK EAVLTMIGVL SESSDGVPKL TVDTNDDLRY LWDYVTTKSY ISALNWFKNE FFVDEWNIAD V VANSENNY FTMASERDVD GTFKLIELAK ASKESLKIIP LGSIENLNYA QEEISKSKIE GGWILLQNIQ MSLSWVKTYL HK HVEETKA AEEHEKFKMF MTCHLTGDKL PAPLLQRTDR VVYEDIPGIL DTVKDLWGSQ FFTGKISGVW SVYCTFLLSW FHA LITART RLVPHGFSKK YYFNDCDFQF ASVYLENVLA TNSTNNIPWA QVRDHIATIV YGGKIDEEKD LEVVAKLCAH VFCG SDNLQ IVPGVRIPQP LLQQSEEEER ARLTAILSNT IEPADSLSSW LQLPRESILD YERLQAKEVA SSTEQLLQEM UniProtKB: Dynein heavy chain, cytoplasmic |
-分子 #2: Nuclear distribution protein PAC1
| 分子 | 名称: Nuclear distribution protein PAC1 / タイプ: protein_or_peptide / ID: 2 / コピー数: 2 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: |
| 分子量 | 理論値: 57.030617 KDa |
| 配列 | 文字列: GMTNWQQQLP LTDTQKNELD KSVLRYLNWN YKQTVRHEHA QDYESVRHAI VTLSGFLLQE SVDRQEFISN NDTSNESMVD IDELLLPKK WNSIVRLQKK IIELEQNTET LVSQIKDLNT QVSELAQFKP TTSNGTSAHN VLKWIPRNLP SCLINVESSV T SVKLHPNL ...文字列: GMTNWQQQLP LTDTQKNELD KSVLRYLNWN YKQTVRHEHA QDYESVRHAI VTLSGFLLQE SVDRQEFISN NDTSNESMVD IDELLLPKK WNSIVRLQKK IIELEQNTET LVSQIKDLNT QVSELAQFKP TTSNGTSAHN VLKWIPRNLP SCLINVESSV T SVKLHPNL PIVFVATDHG KLYAFDLFNY TIPLASLQSH TKAITSMDVL FTNYTNSSKK NYLVIVTASK DLQIHVFKWV SE ECKFQQI RSLLGHEHIV SAVKIWQKNN DVHIASCSRD QTVKIWDFHN GWSLKTFQPH SQWVRSIDVL GDYIISGSHD TTL RLTHWP SGNGLSVGTG HEFPIEKVKF IHFIEDSPEI RFRTPSTDRY KNWGMQYCVS ASRDRTIKIW EIPLPTLMAH RAPI PNPTD SNFRCVLTLK GHLSWVRDIS IRGQYLFSCA DDKSVRCWDL NTGQCLHVWE KLHTGFVNCL DLDVDFDSNV TPRQM MVTG GLDCKSNVFM R UniProtKB: Nuclear distribution protein PAC1 |
-分子 #3: ADENOSINE-5'-TRIPHOSPHATE
| 分子 | 名称: ADENOSINE-5'-TRIPHOSPHATE / タイプ: ligand / ID: 3 / コピー数: 3 / 式: ATP |
|---|---|
| 分子量 | 理論値: 507.181 Da |
| Chemical component information |  ChemComp-ATP: |
-分子 #4: ADENOSINE-5'-DIPHOSPHATE
| 分子 | 名称: ADENOSINE-5'-DIPHOSPHATE / タイプ: ligand / ID: 4 / コピー数: 1 / 式: ADP |
|---|---|
| 分子量 | 理論値: 427.201 Da |
| Chemical component information |  ChemComp-ADP: |
-分子 #5: MAGNESIUM ION
| 分子 | 名称: MAGNESIUM ION / タイプ: ligand / ID: 5 / コピー数: 2 / 式: MG |
|---|---|
| 分子量 | 理論値: 24.305 Da |
-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | particle |
-試料調製
| 緩衝液 | pH: 7.4 |
|---|---|
| グリッド | 詳細: Quantifoil R2/2 grids with gold foil were used. A monolayer of streptavidin crystals was deposited prior to applying the sample. The streptavidin monolayer acted as an affinity surface for ...詳細: Quantifoil R2/2 grids with gold foil were used. A monolayer of streptavidin crystals was deposited prior to applying the sample. The streptavidin monolayer acted as an affinity surface for the biotinylated sample. |
| 凍結 | 凍結剤: ETHANE / チャンバー内湿度: 100 % / チャンバー内温度: 295 K / 装置: FEI VITROBOT MARK II |
| 詳細 | The yeast dynein was biotinylated prior to complex formation. |
- 電子顕微鏡法
電子顕微鏡法
| 顕微鏡 | FEI TITAN KRIOS |
|---|---|
| 温度 | 最低: 70.0 K / 最高: 70.0 K |
| 撮影 | フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 検出モード: SUPER-RESOLUTION / デジタル化 - 画像ごとのフレーム数: 1-50 / 撮影したグリッド数: 1 / 実像数: 2229 / 平均露光時間: 10.0 sec. / 平均電子線量: 58.3 e/Å2 |
| 電子線 | 加速電圧: 300 kV / 電子線源:  FIELD EMISSION GUN FIELD EMISSION GUN |
| 電子光学系 | 照射モード: FLOOD BEAM / 撮影モード: BRIGHT FIELD / Cs: 2.7 mm / 最大 デフォーカス(公称値): 2.7 µm / 最小 デフォーカス(公称値): 2.0 µm |
| 試料ステージ | 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER ホルダー冷却材: NITROGEN |
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |

