MUB1-RAD6-UBR2 ubiquitin ligase complex / RAD6-UBR2 ubiquitin ligase complex / Rad6-Rad18 complex / regulation of dipeptide transport / UBR1-RAD6 ubiquitin ligase complex / sno(s)RNA transcription / proteasome regulatory particle binding / HULC complex / error-free postreplication DNA repair / stress-induced homeostatically regulated protein degradation pathway ...MUB1-RAD6-UBR2 ubiquitin ligase complex / RAD6-UBR2 ubiquitin ligase complex / Rad6-Rad18 complex / regulation of dipeptide transport / UBR1-RAD6 ubiquitin ligase complex / sno(s)RNA transcription / proteasome regulatory particle binding / HULC complex / error-free postreplication DNA repair / stress-induced homeostatically regulated protein degradation pathway / : / meiotic DNA double-strand break formation / ubiquitin-dependent protein catabolic process via the N-end rule pathway / mitochondria-associated ubiquitin-dependent protein catabolic process / cytoplasm protein quality control by the ubiquitin-proteasome system / : / telomere maintenance via recombination / ribosome-associated ubiquitin-dependent protein catabolic process / proteasome regulatory particle, base subcomplex / error-free translesion synthesis / E2 ubiquitin-conjugating enzyme / sporulation resulting in formation of a cellular spore / proteasome binding / ubiquitin conjugating enzyme activity / Antigen processing: Ubiquitination & Proteasome degradation / cellular response to unfolded protein / error-prone translesion synthesis / protein monoubiquitination / ubiquitin ligase complex / subtelomeric heterochromatin formation / mitotic G1 DNA damage checkpoint signaling / ERAD pathway / Maturation of protein E / Maturation of protein E / ER Quality Control Compartment (ERQC) / Myoclonic epilepsy of Lafora / FLT3 signaling by CBL mutants / IRAK2 mediated activation of TAK1 complex / Prevention of phagosomal-lysosomal fusion / Alpha-protein kinase 1 signaling pathway / Glycogen synthesis / IRAK1 recruits IKK complex / IRAK1 recruits IKK complex upon TLR7/8 or 9 stimulation / Endosomal Sorting Complex Required For Transport (ESCRT) / Membrane binding and targetting of GAG proteins / Negative regulation of FLT3 / Regulation of TBK1, IKKε (IKBKE)-mediated activation of IRF3, IRF7 / PTK6 Regulates RTKs and Their Effectors AKT1 and DOK1 / Regulation of TBK1, IKKε-mediated activation of IRF3, IRF7 upon TLR3 ligation / IRAK2 mediated activation of TAK1 complex upon TLR7/8 or 9 stimulation / Constitutive Signaling by NOTCH1 HD Domain Mutants / NOTCH2 Activation and Transmission of Signal to the Nucleus / TICAM1,TRAF6-dependent induction of TAK1 complex / TICAM1-dependent activation of IRF3/IRF7 / APC/C:Cdc20 mediated degradation of Cyclin B / Regulation of FZD by ubiquitination / Downregulation of ERBB4 signaling / APC-Cdc20 mediated degradation of Nek2A / p75NTR recruits signalling complexes / InlA-mediated entry of Listeria monocytogenes into host cells / TRAF6 mediated IRF7 activation in TLR7/8 or 9 signaling / NF-kB is activated and signals survival / TRAF6-mediated induction of TAK1 complex within TLR4 complex / Regulation of pyruvate metabolism / Pexophagy / Regulation of innate immune responses to cytosolic DNA / NRIF signals cell death from the nucleus / Downregulation of ERBB2:ERBB3 signaling / Regulation of PTEN localization / VLDLR internalisation and degradation / Activated NOTCH1 Transmits Signal to the Nucleus / Synthesis of active ubiquitin: roles of E1 and E2 enzymes / Translesion synthesis by REV1 / TICAM1, RIP1-mediated IKK complex recruitment / Regulation of BACH1 activity / Translesion synthesis by POLK / InlB-mediated entry of Listeria monocytogenes into host cell / JNK (c-Jun kinases) phosphorylation and activation mediated by activated human TAK1 / MAP3K8 (TPL2)-dependent MAPK1/3 activation / Activation of IRF3, IRF7 mediated by TBK1, IKKε (IKBKE) / Downregulation of TGF-beta receptor signaling / Translesion synthesis by POLI / Josephin domain DUBs / IKK complex recruitment mediated by RIP1 / Gap-filling DNA repair synthesis and ligation in GG-NER / PINK1-PRKN Mediated Mitophagy / TGF-beta receptor signaling in EMT (epithelial to mesenchymal transition) / Regulation of activated PAK-2p34 by proteasome mediated degradation / TNFR1-induced NF-kappa-B signaling pathway / TCF dependent signaling in response to WNT / Regulation of NF-kappa B signaling / activated TAK1 mediates p38 MAPK activation / Autodegradation of Cdh1 by Cdh1:APC/C / APC/C:Cdc20 mediated degradation of Securin / Asymmetric localization of PCP proteins / Regulation of signaling by CBL / N-glycan trimming in the ER and Calnexin/Calreticulin cycle / Ubiquitin-dependent degradation of Cyclin D / NOTCH3 Activation and Transmission of Signal to the Nucleus / Negative regulators of DDX58/IFIH1 signaling 類似検索 - 分子機能
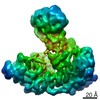
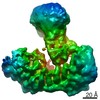
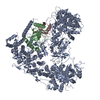
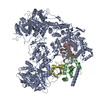
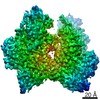
ジャーナル: Nature / 年: 2021 タイトル: Structural insights into Ubr1-mediated N-degron polyubiquitination. 著者: Man Pan / Qingyun Zheng / Tian Wang / Lujun Liang / Junxiong Mao / Chong Zuo / Ruichao Ding / Huasong Ai / Yuan Xie / Dong Si / Yuanyuan Yu / Lei Liu / Minglei Zhao / 要旨: The N-degron pathway targets proteins that bear a destabilizing residue at the N terminus for proteasome-dependent degradation. In yeast, Ubr1-a single-subunit E3 ligase-is responsible for the Arg/N- ...The N-degron pathway targets proteins that bear a destabilizing residue at the N terminus for proteasome-dependent degradation. In yeast, Ubr1-a single-subunit E3 ligase-is responsible for the Arg/N-degron pathway. How Ubr1 mediates the initiation of ubiquitination and the elongation of the ubiquitin chain in a linkage-specific manner through a single E2 ubiquitin-conjugating enzyme (Ubc2) remains unknown. Here we developed chemical strategies to mimic the reaction intermediates of the first and second ubiquitin transfer steps, and determined the cryo-electron microscopy structures of Ubr1 in complex with Ubc2, ubiquitin and two N-degron peptides, representing the initiation and elongation steps of ubiquitination. Key structural elements, including a Ubc2-binding region and an acceptor ubiquitin-binding loop on Ubr1, were identified and characterized. These structures provide mechanistic insights into the initiation and elongation of ubiquitination catalysed by Ubr1.
EMPIAR-10886 (タイトル: Single-particle cryoEM data of yeast Ubr1-Ubc2-Ub-N-degron complex (initiation) Data size: 5.7 TB Data #1: Unaligned movie data of yeast Ubr1-Ubc2-Ub-N-degron complex (initiation complex) [micrographs - multiframe])
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 マップデータ
マップデータ 試料
試料 キーワード
キーワード 機能・相同性情報
機能・相同性情報
 Homo sapiens (ヒト) /
Homo sapiens (ヒト) /  データ登録者
データ登録者 引用
引用

 構造の表示
構造の表示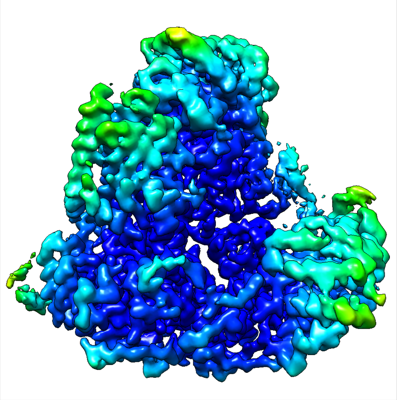
 ダウンロードとリンク
ダウンロードとリンク emd_23806.png
emd_23806.png http://ftp.pdbj.org/pub/emdb/structures/EMD-23806
http://ftp.pdbj.org/pub/emdb/structures/EMD-23806
























 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)































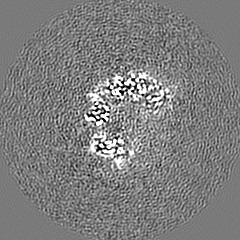



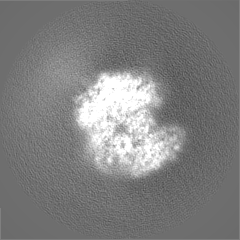







 試料の構成要素
試料の構成要素 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN