 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 |  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
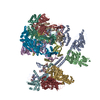
| タイトル | Human Tip60 complex | |||||||||
 マップデータ マップデータ | Overall structure | |||||||||
 試料 試料 |
| |||||||||
 キーワード キーワード | Eukaryotic transcription / Histone acetyltransferase / chromatin remodeling / Complex / TRANSCRIPTION | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報piccolo histone acetyltransferase complex / promoter-enhancer loop anchoring activity / positive regulation of norepinephrine uptake / telomerase RNA localization to Cajal body / regulation of DNA strand elongation / positive regulation of telomere maintenance in response to DNA damage / sperm DNA condensation / histone chaperone activity / establishment of protein localization to chromatin / cellular response to cytochalasin B ...piccolo histone acetyltransferase complex / promoter-enhancer loop anchoring activity / positive regulation of norepinephrine uptake / telomerase RNA localization to Cajal body / regulation of DNA strand elongation / positive regulation of telomere maintenance in response to DNA damage / sperm DNA condensation / histone chaperone activity / establishment of protein localization to chromatin / cellular response to cytochalasin B / bBAF complex / npBAF complex / R2TP complex / nBAF complex / brahma complex / regulation of transepithelial transport / dynein axonemal particle / neural retina development / morphogenesis of a polarized epithelium / structural constituent of postsynaptic actin cytoskeleton / GBAF complex / Formation of annular gap junctions / Formation of the dystrophin-glycoprotein complex (DGC) / protein localization to adherens junction / Gap junction degradation / regulation of G0 to G1 transition / Folding of actin by CCT/TriC / Swr1 complex / dense body / Cell-extracellular matrix interactions / postsynaptic actin cytoskeleton / Tat protein binding / RPAP3/R2TP/prefoldin-like complex / protein antigen binding / chromatin-protein adaptor activity / Prefoldin mediated transfer of substrate to CCT/TriC / RSC-type complex / Ino80 complex / blastocyst formation / regulation of nucleotide-excision repair / regulation of double-strand break repair / adherens junction assembly / RHOF GTPase cycle / Adherens junctions interactions / apical protein localization / Sensory processing of sound by outer hair cells of the cochlea / tight junction / box C/D snoRNP assembly / Interaction between L1 and Ankyrins / SWI/SNF complex / regulation of mitotic metaphase/anaphase transition / protein folding chaperone complex / Sensory processing of sound by inner hair cells of the cochlea / positive regulation of T cell differentiation / regulation of norepinephrine uptake / apical junction complex / transporter regulator activity / Formation of Senescence-Associated Heterochromatin Foci (SAHF) / positive regulation of double-strand break repair / spinal cord development / maintenance of blood-brain barrier / nitric-oxide synthase binding / negative regulation of gene expression, epigenetic / establishment or maintenance of cell polarity / cortical cytoskeleton / NuA4 histone acetyltransferase complex / regulation of chromosome organization / positive regulation of stem cell population maintenance / RUNX1 interacts with co-factors whose precise effect on RUNX1 targets is not known / Transcriptional Regulation by E2F6 / Regulation of MITF-M-dependent genes involved in pigmentation / Recycling pathway of L1 / brush border / regulation of G1/S transition of mitotic cell cycle / TFIID-class transcription factor complex binding / regulation of DNA replication / MLL1 complex / kinesin binding / regulation of embryonic development / Telomere Extension By Telomerase / EPH-ephrin mediated repulsion of cells / negative regulation of cell differentiation / somatic stem cell population maintenance / spermatid development / regulation of synaptic vesicle endocytosis / RHO GTPases Activate WASPs and WAVEs / enzyme-substrate adaptor activity / positive regulation of myoblast differentiation / RHO GTPases activate IQGAPs / regulation of protein localization to plasma membrane / positive regulation of double-strand break repair via homologous recombination / RNA polymerase II core promoter sequence-specific DNA binding / regulation of DNA repair / cytoskeleton organization / EPHB-mediated forward signaling / Deposition of new CENPA-containing nucleosomes at the centromere / substantia nigra development / telomere maintenance / axonogenesis / DNA helicase activity 類似検索 - 分子機能 | |||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | |||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 2.52 Å | |||||||||
 データ登録者 データ登録者 | Li C / Smirnova E / Schnitzler C / Crucifix C / Concordet JP / Brion A / Poterszman A / Schultz P / Papai G / Ben-Shem A | |||||||||
| 資金援助 |  フランス, 1件 フランス, 1件
| |||||||||
 引用 引用 | ジャーナル: Nature / 年: 2024 タイトル: Structure of the human TIP60-C histone exchange and acetyltransferase complex. 著者: Changqing Li / Ekaterina Smirnova / Charlotte Schnitzler / Corinne Crucifix / Jean Paul Concordet / Alice Brion / Arnaud Poterszman / Patrick Schultz / Gabor Papai / Adam Ben-Shem / 要旨: Chromatin structure is a key regulator of DNA transcription, replication and repair. In humans, the TIP60-EP400 complex (TIP60-C) is a 20-subunit assembly that affects chromatin structure through two ...Chromatin structure is a key regulator of DNA transcription, replication and repair. In humans, the TIP60-EP400 complex (TIP60-C) is a 20-subunit assembly that affects chromatin structure through two enzymatic activities: ATP-dependent exchange of histone H2A-H2B for H2A.Z-H2B, and histone acetylation. In yeast, however, these activities are performed by two independent complexes-SWR1 and NuA4, respectively. How the activities of the two complexes are merged into one supercomplex in humans, and what this association entails for the structure and mechanism of the proteins and their recruitment to chromatin, are unknown. Here we describe the structure of the endogenous human TIP60-C. We find a three-lobed architecture composed of SWR1-like (SWR1L) and NuA4-like (NuA4L) parts, which associate with a TRRAP activator-binding module. The huge EP400 subunit contains the ATPase motor, traverses the junction between SWR1L and NuA4L twice and constitutes the scaffold of the three-lobed architecture. NuA4L is completely rearranged compared with its yeast counterpart. TRRAP is flexibly tethered to NuA4L-in stark contrast to its robust connection to the completely opposite side of NuA4 in yeast. A modelled nucleosome bound to SWR1L, supported by tests of TIP60-C activity, suggests that some aspects of the histone exchange mechanism diverge from what is seen in yeast. Furthermore, a fixed actin module (as opposed to the mobile actin subcomplex in SWR1; ref. ), the flexibility of TRRAP and the weak effect of extranucleosomal DNA on exchange activity lead to a different, activator-based mode of enlisting TIP60-C to chromatin. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 添付画像 |
|---|

- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_18581.map.gz | 616.7 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-18581-v30.xmlemd-18581.xml | 27 KB 27 KB | 表示 表示 | EMDBヘッダ |
| FSC (解像度算出) | emd_18581_fsc.xml | 22.7 KB | 表示 | FSCデータファイル |
| 画像 |  emd_18581.png emd_18581.png | 78.8 KB | ||
| マスクデータ | emd_18581_msk_1.map | 1.2 GB | マスクマップ | |
| Filedesc metadata | emd-18581.cif.gz | 9.2 KB | ||
| その他 | emd_18581_half_map_1.map.gzemd_18581_half_map_2.map.gz | 1.1 GB 1.1 GB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-18581ftp://ftp.pdbj.org/pub/emdb/structures/EMD-18581 http://ftp.pdbj.org/pub/emdb/structures/EMD-18581ftp://ftp.pdbj.org/pub/emdb/structures/EMD-18581 | HTTPS FTP |
-検証レポート
| 文書・要旨 | emd_18581_validation.pdf.gz | 1.1 MB | 表示 | EMDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | emd_18581_full_validation.pdf.gz | 1.1 MB | 表示 | |
| XML形式データ | emd_18581_validation.xml.gz | 30.5 KB | 表示 | |
| CIF形式データ | emd_18581_validation.cif.gz | 39.2 KB | 表示 | |
| アーカイブディレクトリ | https://ftp.pdbj.org/pub/emdb/validation_reports/EMD-18581ftp://ftp.pdbj.org/pub/emdb/validation_reports/EMD-18581 | HTTPS FTP |
-関連構造データ









-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| 「今月の分子」の関連する項目 |




























-マップ
| ファイル | ダウンロード / ファイル: emd_18581.map.gz / 形式: CCP4 / 大きさ: 1.2 GB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | Overall structure | ||||||||||||||||||||||||||||||||||||
| 投影像・断面図 | 画像のコントロール
画像は Spider により作成 | ||||||||||||||||||||||||||||||||||||
| ボクセルのサイズ | X=Y=Z: 0.73 Å | ||||||||||||||||||||||||||||||||||||
| 密度 |
| ||||||||||||||||||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||||||||||||||||||
| 詳細 | EMDB XML:
|
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-添付データ
-マスク #1
| ファイル | emd_18581_msk_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |








-ハーフマップ: Half map A
| ファイル | emd_18581_half_map_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | Half map A | ||||||||||||
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |






-ハーフマップ: Half map B
| ファイル | emd_18581_half_map_2.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | Half map B | ||||||||||||
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |








- 試料の構成要素
試料の構成要素
-全体 : Tip60 complex
| 全体 | 名称: Tip60 complex |
|---|---|
| 要素 |
|
-超分子 #1: Tip60 complex
| 超分子 | 名称: Tip60 complex / タイプ: cell / ID: 1 / 親要素: 0 / 含まれる分子: all |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) / 株: K562 |
-分子 #1: EP400
| 分子 | 名称: EP400 / タイプ: protein_or_peptide / ID: 1 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 配列 | 文字列: MHHGTGPQNV QHQLQRSRAC PGSEGEEQPA HPNPPPSPAA PFAPSASPSA PQSPSYQIQQ LMNRSPATG QNVNITLQSV GPVVGGNQQI TLAPLPLPSP TSPGFQFSAQ PRRFEHGSPS Y IQVTSPLS QQVQTQSPTQ PSPGPGQALQ NVRAGAPGPG LGLCSSSPTG ...文字列: MHHGTGPQNV QHQLQRSRAC PGSEGEEQPA HPNPPPSPAA PFAPSASPSA PQSPSYQIQQ LMNRSPATG QNVNITLQSV GPVVGGNQQI TLAPLPLPSP TSPGFQFSAQ PRRFEHGSPS Y IQVTSPLS QQVQTQSPTQ PSPGPGQALQ NVRAGAPGPG LGLCSSSPTG GFVDASVLVR QI SLSPSSG GHFVFQDGSG LTQIAQGAQV QLQHPGTPIT VRERRPSQPH TQSGGTIHHL GPQ SPAAAG GAGLQPLASP SHITTANLPP QISSIIQGQL VQQQQVLQGP PLPRPLGFER TPGV LLPGA GGAAGFGMTS PPPPTSPSRT AVPPGLSSLP LTSVGNTGMK KVPKKLEEIP PASPE MAQM RKQCLDYHYQ EMQALKEVFK EYLIELFFLQ HFQGNMMDFL AFKKKHYAPL QAYLRQ NDL DIEEEEEEEE EEEEKSEVIN DEVKVVTGKD GQTGTPVAIA TQLPPKVSAA FSSQQQP FQ QALAGSLVAG AGSTVETDLF KRQQAMPSTG MAEQSKRPRL EVGHQGVVFQ HPGADAGV P LQQLMPTAQG GMPPTPQAAQ LAGQRQSQQQ YDPSTGPPVQ NAASLHTPLP QLPGRLPPA GVPTAALSSA LQFAQQPQVV EAQTQLQIPV KTQQPNVPIP APPSSQLPIP PSQPAQLALH VPTPGKVQV QASQLSSLPQ MVASTRLPVD PAPPCPRPLP TSSTSSLAPV SGSGPGPSPA R SSPVNRPS SATNKALSPV TSRTPGVVAS APTKPQSPAQ NATSSQDSSQ DTLTEQITLE NQ VHQRIAE LRKAGLWSQR RLPKLQEAPR PKSHWDYLLE EMQWMATDFA QERRWKVAAA KKL VRTVVR HHEEKQLREE RGKKEEQSRL RRIAASTARE IECFWSNIEQ VVEIKLRVEL EEKR KKALN LQKVSRRGKE LRPKGFDALQ ESSLDSGMSG RKRKASISLT DDEVDDEEET IEEEE ANEG VVDHQTELSN LAKEAELPLL DLMKLYEGAF LPSSQWPRPK PDGEDTSGEE DADDCP GDR ESRKDLVLID SLFIMDQFKA AERMNIGKPN AKDIADVTAV AEAILPKGSA RVTTSVK FN APSLLYGALR DYQKIGLDWL AKLYRKNLNG ILADEAGLGK TVQIIAFFAH LACNEGNW G PHLVVVRSCN ILKWELELKR WCPGLKILSY IGSHRELKAK RQEWAEPNSF HVCITSYTQ FFRGLTAFTR VRWKCLVIDE MQRVKGMTER HWEAVFTLQS QQRLLLIDSP LHNTFLELWT MVHFLVPGI SRPYLSSPLR APSEESQDYY HKVVIRLHRV TQPFILRRTK RDVEKQLTKK Y EHVLKCRL SNRQKALYED VILQPGTQEA LKSGHFVNVL SILVRLQRIC NHPGLVEPRH PG SSYVAGP LEYPSASLIL KALERDFWKE ADLSMFDLIG LENKITRHEA ELLSKKKIPR KLM EEISTS AAPAARPAAA KLKASRLFQP VQYGQKPEGR TVAFPSTHPP RTAAPTTASA APQG PLRGR PPIATFSANP EAKAAAAPFQ TSQASASAPR HQPASASSTA ASPAHPAKLR AQTTA QAST PGQPPPQPQA PSHAAGQSAL PQRLVLPSQA QARLPSGEVV KIAQLASITG PQSRVA QPE TPVTLQFQGS KFTLSHSQLR QLTAGQPLQL QGSVLQIVSA PGQPYLRAPG PVVMQTV SQ AGAVHGALGS KPPAGGPSPA PLTPQVGVPG RVAVNALAVG EPGTASKPAS PIGGPTQE E KTRLLKERLD QIYLVNERRC SQAPVYGRDL LRICALPSHG RVQWRGSLDG RRGKEAGPA HSYTSSSESP SELMLTLCRC GESLQDVIDR VAFVIPPVVA APPSLRVPRP PPLYSHRMRI LRQGLREHA APYFQQLRQT TAPRLLQFPE LRLVQFDSGK LEALAILLQK LKSEGRRVLI L SQMILMLD ILEMFLNFHY LTYVRIDENA SSEQRQELMR SFNRDRRIFC AILSTHSRTT GI NLVEADT VVFYDNDLNP VMDAKAQEWC DRIGRCKDIH IYRLVSGNSI EEKLLKNGTK DLI REVAAQ GNDYSMAFLT QRTIQELFEV YSPMDDAGFP VKAEEFVVLS QEPSVTETIA PKIA RPFIE ALKSIEYLEE DAQKSAQEGV LGPHTDALSS DSENMPCDEE PSQLEELADF MEQLT PIEK YALNYLELFH TSIEQEKERN SEDAVMTAVR AWEFWNLKTL QEREARLRLE QEEAEL LTY TREDAYSMEY VYEDVDGQTE VMPLWTPPTP PQDDSDIYLD SVMCLMYEAT PIPEAKL PP VYVRKERKRH KTDPSAAGRK KKQRHGEAVV PPRSLFDRAT PGLLKIRREG KEQKKNIL L KQQVPFAKPL PTFAKPTAEP GQDNPEWLIS EDWALLQAVK QLLELPLNLT IVSPAHTPN WDLVSDVVNS CSRIYRSSKQ CRNRYENVII PREEGKSKNN RPLRTSQIYA QDENATHTQL YTSHFDLMK MTAGKRSPPI KPLLGMNPFQ KNPKHASVLA ESGINYDKPL PPIQVASLRA E RIAKEKKA LADQQKAQQP AVAQPPPPQP QPPPPPQQPP PPLPQPQAAG SQPPAGPPAV QP QPQPQPQ TQPQPVQAPA KAQPAITTGG SAAVLAGTIK TSVTGTSMPT GAVSGNVIVN TIA GVPAAT FQSINKRLAS PVAPGALTTP GGSAPAQVVH TQPPPRAVGS PATATPDLVS MATT QGVRA VTSVTASAVV TTNLTPVQTP ARSLVPQVSQ ATGVQLPGKT ITPAHFQLLR QQQQQ QQQQ QQQQQQQQQQ QQQQQQQQQQ TTTTSQVQVP QIQGQAQSPA QIKAVGKLTP EHLIKM QKQ KLQMPPQPPP PQAQSAPPQP TAQVQVQTSQ PPQQQSPQLT TVTAPRPGAL LTGTTVA NL QVARLTRVPT SQLQAQGQMQ TQAPQPAQVA LAKPPVVSVP AAVVSSPGVT TLPMNVAG I SVAIGQPQKA AGQTVVAQPV HMQQLLKLKQ QAVQQQKAIQ PQAAQGPAAV QQKITAQQI TTPGAQQKVA YAAQPALKTQ FLTTPISQAQ KLAGAQQVQT QIQVAKLPQV VQQQTPVASI QQVASASQQ ASPQTVALTQ ATAAGQQVQM IPAVTATAQV VQQKLIQQQV VTTASAPLQT P GAPNPAQV PASSDSPSQQ PKLQMRVPAV RLKTPTKPPC Q UniProtKB: E1A-binding protein p400 |
-分子 #2: EPC1
| 分子 | 名称: EPC1 / タイプ: protein_or_peptide / ID: 2 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 配列 | 文字列: MSKLSFRARA LDASKPLPVF RCEDLPDLHE YASINRAVPQ MPTGMEKEEE SEHHLQRAIS AQQVYGEKRD NMVIPVPEAE SNIAYYESIY PGEFKMPKQL IHIQPFSLDA EQPDYDLDSE DEVFVNKLKK KMDICPLQFE EMIDRLEKGS GQQPVSLQEA KLLLKEDDEL ...文字列: MSKLSFRARA LDASKPLPVF RCEDLPDLHE YASINRAVPQ MPTGMEKEEE SEHHLQRAIS AQQVYGEKRD NMVIPVPEAE SNIAYYESIY PGEFKMPKQL IHIQPFSLDA EQPDYDLDSE DEVFVNKLKK KMDICPLQFE EMIDRLEKGS GQQPVSLQEA KLLLKEDDEL IREVYEYWIK KRKNCRGPSL IPSVKQEKRD GSSTNDPYVA FRRRTEKMQT RKNRKNDEAS YEKMLKLRRD LSRAVTILEM IKRREKSKRE LLHLTLEIME KRYNLGDYNG EIMSEVMAQR QPMKPTYAIP IIPITNSSQF KHQEAMDVKE FKVNKQDKAD LIRPKRKYEK KPKVLPSSAA ATPQQTSPAA LPVFNAKDLN QYDFPSSDEE PLSQVLSGSS EAEEDNDPDG PFAFRRKAGC QYYAPHLDQT GNWPWTSPKD GGLGDVRYRY CLTTLTVPQR CIGFARRRVG RGGRVLLDRA HSDYDSVFHH LDLEMLSSPQ HSPVNQFANT SETNTSDKSF SKDLSQILVN IKSCRWRHFR PRTPSLHDSD NDELSCRKLY RSINRTGTAQ PGTQTCSTST QSKSSSGSAH FAFTAEQYQQ HQQQLALMQK QQLAQIQQQQ ANSNSSTNTS QNLASNQQKS GFRLNIQGLE RTLQGFVSKT LDSASAQFAA SALVTSEQLM GFKMKDDVVL GIGVNGVLPA SGVYKGLHLS STTPTALVHT SPSTAGSALL QPSNITQTSS SHSALSHQVT AANSATTQVL IGNNIRLTVP SSVATVNSIA PINARHIPRT LSAVPSSALK LAAAANCQVS KVPSSSSVDS VPRENHESEK PALNNIADNT VAMEVT UniProtKB: Enhancer of polycomb homolog 1 |
-分子 #3: VPS72
| 分子 | 名称: VPS72 / タイプ: protein_or_peptide / ID: 3 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 配列 | 文字列: MSLAGGRAPR KTAGNRLSGL LEAEEEDEFY QTTYGGFTEE SGDDEYQGDQ SDTEDEVDSD FDIDEGDEPS SDGEAEEPRR KRRVVTKAYK EPLKSLRPRK VNTPAGSSQK AREEKALLPL ELQDDGSDSR KSMRQSTAEH TRQTFLRVQE RQGQSRRRKG PHCERPLTQE ...文字列: MSLAGGRAPR KTAGNRLSGL LEAEEEDEFY QTTYGGFTEE SGDDEYQGDQ SDTEDEVDSD FDIDEGDEPS SDGEAEEPRR KRRVVTKAYK EPLKSLRPRK VNTPAGSSQK AREEKALLPL ELQDDGSDSR KSMRQSTAEH TRQTFLRVQE RQGQSRRRKG PHCERPLTQE ELLREAKITE ELNLRSLETY ERLEADKKKQ VHKKRKCPGP IITYHSVTVP LVGEPGPKEE NVDIEGLDPA PSVSALTPHA GTGPVNPPAR CSRTFITFSD DATFEEWFPQ GRPPKVPVRE VCPVTHRPAL YRDPVTDIPY ATARAFKIIR EAYKKYITAH GLPPTASALG PGPPPPEPLP GSGPRALRQK IVIK UniProtKB: Vacuolar protein sorting-associated protein 72 homolog |
-分子 #4: DMAP1
| 分子 | 名称: DMAP1 / タイプ: protein_or_peptide / ID: 4 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 配列 | 文字列: MATGADVRDI LELGGPEGDA ASGTISKKDI INPDKKKSKK SSETLTFKRP EGMHREVYAL LYSDKKDAPP LLPSDTGQGY RTVKAKLGSK KVRPWKWMPF TNPARKDGAM FFHWRRAAEE GKDYPFARFN KTVQVPVYSE QEYQLYLHDD AWTKAETDHL FDLSRRFDLR ...文字列: MATGADVRDI LELGGPEGDA ASGTISKKDI INPDKKKSKK SSETLTFKRP EGMHREVYAL LYSDKKDAPP LLPSDTGQGY RTVKAKLGSK KVRPWKWMPF TNPARKDGAM FFHWRRAAEE GKDYPFARFN KTVQVPVYSE QEYQLYLHDD AWTKAETDHL FDLSRRFDLR FVVIHDRYDH QQFKKRSVED LKERYYHICA KLANVRAVPG TDLKIPVFDA GHERRRKEQL ERLYNRTPEQ VAEEEYLLQE LRKIEARKKE REKRSQDLQK LITAADTTAE QRRTERKAPK KKLPQKKEAE KPAVPETAGI KFPDFKSAGV TLRSQRMKLP SSVGQKKIKA LEQMLLELGV ELSPTPTEEL VHMFNELRSD LVLLYELKQA CANCEYELQM LRHRHEALAR AGVLGGPATP ASGPGPASAE PAVTEPGLGP DPKDTIIDVV GAPLTPNSRK RRESASSSSS VKKAKKP UniProtKB: DNA methyltransferase 1-associated protein 1 |
-分子 #5: ACL6A
| 分子 | 名称: ACL6A / タイプ: protein_or_peptide / ID: 5 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 配列 | 文字列: MSGGVYGGDE VGALVFDIGS YTVRAGYAGE DCPKVDFPTA IGMVVERDDG STLMEIDGDK GKQGGPTYYI DTNALRVPRE NMEAISPLKN GMVEDWDSFQ AILDHTYKMH VKSEASLHPV LMSEAPWNTR AKREKLTELM FEHYNIPAFF LCKTAVLTAF ANGRSTGLIL ...文字列: MSGGVYGGDE VGALVFDIGS YTVRAGYAGE DCPKVDFPTA IGMVVERDDG STLMEIDGDK GKQGGPTYYI DTNALRVPRE NMEAISPLKN GMVEDWDSFQ AILDHTYKMH VKSEASLHPV LMSEAPWNTR AKREKLTELM FEHYNIPAFF LCKTAVLTAF ANGRSTGLIL DSGATHTTAI PVHDGYVLQQ GIVKSPLAGD FITMQCRELF QEMNIELVPP YMIASKEAVR EGSPANWKRK EKLPQVTRSW HNYMCNCVIQ DFQASVLQVS DSTYDEQVAA QMPTVHYEFP NGYNCDFGAE RLKIPEGLFD PSNVKGLSGN TMLGVSHVVT TSVGMCDIDI RPGLYGSVIV AGGNTLIQSF TDRLNRELSQ KTPPSMRLKL IANNTTVERR FSSWIGGSIL ASLGTFQQMW ISKQEYEEGG KQCVERKCP UniProtKB: Actin-like protein 6A |
-分子 #6: ACTIN
| 分子 | 名称: ACTIN / タイプ: protein_or_peptide / ID: 6 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 配列 | 文字列: MDDDIAALVV DNGSGMCKAG FAGDDAPRAV FPSIVGRPRH QGVMVGMGQK DSYVGDEAQS KRGILTLKYP IEHGIVTNWD DMEKIWHHTF YNELRVAPEE HPVLLTEAPL NPKANREKMT QIMFETFNTP AMYVAIQAVL SLYASGRTTG IVMDSGDGVT HTVPIYEGYA ...文字列: MDDDIAALVV DNGSGMCKAG FAGDDAPRAV FPSIVGRPRH QGVMVGMGQK DSYVGDEAQS KRGILTLKYP IEHGIVTNWD DMEKIWHHTF YNELRVAPEE HPVLLTEAPL NPKANREKMT QIMFETFNTP AMYVAIQAVL SLYASGRTTG IVMDSGDGVT HTVPIYEGYA LPHAILRLDL AGRDLTDYLM KILTERGYSF TTTAEREIVR DIKEKLCYVA LDFEQEMATA ASSSSLEKSY ELPDGQVITI GNERFRCPEA LFQPSFLGME SCGIHETTFN SIMKCDVDIR KDLYANTVLS GGTTMYPGIA DRMQKEITAL APSTMKIKII APPERKYSVW IGGSILASLS TFQQMWISKQ EYDESGPSIV HRKCF UniProtKB: Actin, cytoplasmic 1 |
-分子 #7: RUVBL1
| 分子 | 名称: RUVBL1 / タイプ: protein_or_peptide / ID: 7 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 配列 | 文字列: MKIEEVKSTT KTQRIASHSH VKGLGLDESG LAKQAASGLV GQENAREACG VIVELIKSKK MAGRAVLLAG PPGTGKTALA LAIAQELGSK VPFCPMVGSE VYSTEIKKTE VLMENFRRAI GLRIKETKEV YEGEVTELTP CETENPMGGY GKTISHVIIG LKTAKGTKQL ...文字列: MKIEEVKSTT KTQRIASHSH VKGLGLDESG LAKQAASGLV GQENAREACG VIVELIKSKK MAGRAVLLAG PPGTGKTALA LAIAQELGSK VPFCPMVGSE VYSTEIKKTE VLMENFRRAI GLRIKETKEV YEGEVTELTP CETENPMGGY GKTISHVIIG LKTAKGTKQL KLDPSIFESL QKERVEAGDV IYIEANSGAV KRQGRCDTYA TEFDLEAEEY VPLPKGDVHK KKEIIQDVTL HDLDVANARP QGGQDILSMM GQLMKPKKTE ITDKLRGEIN KVVNKYIDQG IAELVPGVLF VDEVHMLDIE CFTYLHRALE SSIAPIVIFA SNRGNCVIRG TEDITSPHGI PLDLLDRVMI IRTMLYTPQE MKQIIKIRAQ TEGINISEEA LNHLGEIGTK TTLRYSVQLL TPANLLAKIN GKDSIEKEHV EEISELFYDA KSSAKILADQ QDKYMK UniProtKB: RuvB-like 1 |
-分子 #8: RUVBL2
| 分子 | 名称: RUVBL2 / タイプ: protein_or_peptide / ID: 8 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 配列 | 文字列: MATVTATTKV PEIRDVTRIE RIGAHSHIRG LGLDDALEPR QASQGMVGQL AARRAAGVVL EMIREGKIAG RAVLIAGQPG TGKTAIAMGM AQALGPDTPF TAIAGSEIFS LEMSKTEALT QAFRRSIGVR IKEETEIIEG EVVEIQIDRP ATGTGSKVGK LTLKTTEMET ...文字列: MATVTATTKV PEIRDVTRIE RIGAHSHIRG LGLDDALEPR QASQGMVGQL AARRAAGVVL EMIREGKIAG RAVLIAGQPG TGKTAIAMGM AQALGPDTPF TAIAGSEIFS LEMSKTEALT QAFRRSIGVR IKEETEIIEG EVVEIQIDRP ATGTGSKVGK LTLKTTEMET IYDLGTKMIE SLTKDKVQAG DVITIDKATG KISKLGRSFT RARDYDAMGS QTKFVQCPDG ELQKRKEVVH TVSLHEIDVI NSRTQGFLAL FSGDTGEIKS EVREQINAKV AEWREEGKAE IIPGVLFIDE VHMLDIESFS FLNRALESDM APVLIMATNR GITRIRGTSY QSPHGIPIDL LDRLLIVSTT PYSEKDTKQI LRIRCEEEDV EMSEDAYTVL TRIGLETSLR YAIQLITAAS LVCRKRKGTE VQVDDIKRVY SLFLDESRST QYMKEYQDAF LFNELKGETM DTS UniProtKB: RuvB-like 2 |
-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | particle |
-試料調製
| 濃度 | 0.2 mg/mL | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 緩衝液 | pH: 8 構成要素:
| ||||||||||||||||
| グリッド | モデル: UltrAuFoil R1.2/1.3 / 材質: GOLD / メッシュ: 300 / 前処理 - タイプ: PLASMA CLEANING / 前処理 - 時間: 90 sec. | ||||||||||||||||
| 凍結 | 凍結剤: ETHANE / チャンバー内湿度: 95 % / チャンバー内温度: 279 K / 装置: FEI VITROBOT MARK IV |
- 電子顕微鏡法
電子顕微鏡法
| 顕微鏡 | FEI TITAN KRIOS |
|---|---|
| 撮影 | フィルム・検出器のモデル: FEI FALCON IV (4k x 4k) 平均電子線量: 40.0 e/Å2 |
| 電子線 | 加速電圧: 300 kV / 電子線源:  FIELD EMISSION GUN FIELD EMISSION GUN |
| 電子光学系 | 照射モード: FLOOD BEAM / 撮影モード: BRIGHT FIELD / Cs: 2.7 mm 最大 デフォーカス(公称値): 2.8000000000000003 µm 最小 デフォーカス(公称値): 1.2 µm |
| 試料ステージ | 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER ホルダー冷却材: NITROGEN |
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |

