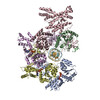
Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-8s0c: H. sapiens ORC1-5 bound to double stranded DNA as part of the MCM... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8s0c | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | H. sapiens ORC1-5 bound to double stranded DNA as part of the MCM-ORC complex | |||||||||||||||
 Components Components |
| |||||||||||||||
 Keywords Keywords | REPLICATION / AAA+ ATPase / DNA helicase | |||||||||||||||
| Function / homology |  Function and homology information Function and homology informationCDC6 association with the ORC:origin complex / polar body extrusion after meiotic divisions / origin recognition complex / E2F-enabled inhibition of pre-replication complex formation / inner kinetochore / nuclear origin of replication recognition complex / nuclear pre-replicative complex / DNA replication preinitiation complex / mitotic DNA replication checkpoint signaling / G1/S-Specific Transcription ...CDC6 association with the ORC:origin complex / polar body extrusion after meiotic divisions / origin recognition complex / E2F-enabled inhibition of pre-replication complex formation / inner kinetochore / nuclear origin of replication recognition complex / nuclear pre-replicative complex / DNA replication preinitiation complex / mitotic DNA replication checkpoint signaling / G1/S-Specific Transcription / regulation of DNA replication / DNA replication origin binding / protein polymerization / Activation of the pre-replicative complex / DNA replication initiation / Activation of ATR in response to replication stress / heterochromatin / Assembly of the ORC complex at the origin of replication / Assembly of the pre-replicative complex / Orc1 removal from chromatin / chromosome, telomeric region / DNA replication / nucleotide binding / centrosome / chromatin binding / nucleolus / chromatin / negative regulation of transcription by RNA polymerase II / ATP hydrolysis activity / DNA binding / nucleoplasm / ATP binding / membrane / metal ion binding / nucleus / cytosol / cytoplasm Similarity search - Function | |||||||||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human)synthetic construct (others) | |||||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 4 Å | |||||||||||||||
 Authors Authors | Greiwe, J.F. / Weissmann, F. / Diffley, J.F.X. / Costa, A. | |||||||||||||||
| Funding support |  United Kingdom, European Union, 4items United Kingdom, European Union, 4items
| |||||||||||||||
 Citation Citation | Journal: Nature / Year: 2024 Title: MCM double hexamer loading visualized with human proteins. Authors: Florian Weissmann / Julia F Greiwe / Thomas Pühringer / Evelyn L Eastwood / Emma C Couves / Thomas C R Miller / John F X Diffley / Alessandro Costa /  Abstract: Eukaryotic DNA replication begins with the loading of the MCM replicative DNA helicase as a head-to-head double hexamer at origins of DNA replication. Our current understanding of how the double ...Eukaryotic DNA replication begins with the loading of the MCM replicative DNA helicase as a head-to-head double hexamer at origins of DNA replication. Our current understanding of how the double hexamer is assembled by the origin recognition complex (ORC), CDC6 and CDT1 comes mostly from budding yeast. Here we characterize human double hexamer (hDH) loading using biochemical reconstitution and cryo-electron microscopy with purified proteins. We show that the human double hexamer engages DNA differently from the yeast double hexamer (yDH), and generates approximately five base pairs of underwound DNA at the interface between hexamers, as seen in hDH isolated from cells. We identify several differences from the yeast double hexamer in the order of factor recruitment and dependencies during hDH assembly. Unlike in yeast, the ORC6 subunit of the ORC is not essential for initial MCM recruitment or hDH loading, but contributes to an alternative hDH assembly pathway that requires an intrinsically disordered region in ORC1, which may work through a MCM-ORC intermediate. Our work presents a detailed view of how double hexamers are assembled in an organism that uses sequence-independent replication origins, provides further evidence for diversity in eukaryotic double hexamer assembly mechanisms, and represents a first step towards reconstitution of DNA replication initiation with purified human proteins. #1: Journal: Biorxiv / Year: 2024Title: MCM Double Hexamer Loading Visualised with Human Proteins Authors: Weissmann, F. / Greiwe, J.F. / Puhringer, T. / Miller, T.C.R. / Diffley, J.F.X. / Costa, A. #2: Journal: Acta Crystallogr D Struct Biol / Year: 2019 Title: Macromolecular structure determination using X-rays, neutrons and electrons: recent developments in Phenix. Authors: Dorothee Liebschner / Pavel V Afonine / Matthew L Baker / Gábor Bunkóczi / Vincent B Chen / Tristan I Croll / Bradley Hintze / Li Wei Hung / Swati Jain / Airlie J McCoy / Nigel W Moriarty ...Authors: Dorothee Liebschner / Pavel V Afonine / Matthew L Baker / Gábor Bunkóczi / Vincent B Chen / Tristan I Croll / Bradley Hintze / Li Wei Hung / Swati Jain / Airlie J McCoy / Nigel W Moriarty / Robert D Oeffner / Billy K Poon / Michael G Prisant / Randy J Read / Jane S Richardson / David C Richardson / Massimo D Sammito / Oleg V Sobolev / Duncan H Stockwell / Thomas C Terwilliger / Alexandre G Urzhumtsev / Lizbeth L Videau / Christopher J Williams / Paul D Adams /   Abstract: Diffraction (X-ray, neutron and electron) and electron cryo-microscopy are powerful methods to determine three-dimensional macromolecular structures, which are required to understand biological ...Diffraction (X-ray, neutron and electron) and electron cryo-microscopy are powerful methods to determine three-dimensional macromolecular structures, which are required to understand biological processes and to develop new therapeutics against diseases. The overall structure-solution workflow is similar for these techniques, but nuances exist because the properties of the reduced experimental data are different. Software tools for structure determination should therefore be tailored for each method. Phenix is a comprehensive software package for macromolecular structure determination that handles data from any of these techniques. Tasks performed with Phenix include data-quality assessment, map improvement, model building, the validation/rebuilding/refinement cycle and deposition. Each tool caters to the type of experimental data. The design of Phenix emphasizes the automation of procedures, where possible, to minimize repetitive and time-consuming manual tasks, while default parameters are chosen to encourage best practice. A graphical user interface provides access to many command-line features of Phenix and streamlines the transition between programs, project tracking and re-running of previous tasks. | |||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8s0c.cif.gz | 418.9 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8s0c.ent.gz | 265.2 KB | Display | PDB format |
| PDBx/mmJSON format | 8s0c.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/s0/8s0cftp://data.pdbj.org/pub/pdb/validation_reports/s0/8s0c | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  19621MC  8s09C  8s0aC  8s0bC  8s0dC  8s0eC  8s0fC C: citing same article ( M: map data used to model this data |
|---|---|
| Similar structure data |






-Links
 PDBj
PDBj








































- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-Origin recognition complex subunit ... , 4 types, 4 molecules ABDE
| #1: Protein | Mass: 97499.867 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: ORC1, ORC1L, PARC1 / Production host:   Spodoptera frugiperda (fall armyworm) / References: UniProt: Q13415 Spodoptera frugiperda (fall armyworm) / References: UniProt: Q13415 |
|---|---|
| #4: Protein | Mass: 66063.375 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: ORC2, ORC2L / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: Q13416 |
| #6: Protein | Mass: 50443.266 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: ORC4, ORC4L / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: O43929 |
| #7: Protein | Mass: 50349.934 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: ORC5, ORC5L / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: O43913 |
-DNA chain , 2 types, 2 molecules XY
| #2: DNA chain | Mass: 7928.126 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) synthetic construct (others) |
|---|---|
| #3: DNA chain | Mass: 8044.230 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) synthetic construct (others) |
-Protein , 1 types, 1 molecules C
| #5: Protein | Mass: 82436.133 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: ORC3, LATHEO, ORC3L / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: Q9UBD5 |
|---|
-Non-polymers , 2 types, 4 molecules 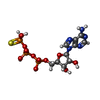

| #8: Chemical |  Mass: 523.247 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H16N5O12P3S / Feature type: SUBJECT OF INVESTIGATION / Comment: ATP-gamma-S, energy-carrying molecule analogue*YM Mass: 523.247 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H16N5O12P3S / Feature type: SUBJECT OF INVESTIGATION / Comment: ATP-gamma-S, energy-carrying molecule analogue*YM#9: Chemical |  Mass: 24.305 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Mg / Feature type: SUBJECT OF INVESTIGATION Mass: 24.305 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Mg / Feature type: SUBJECT OF INVESTIGATION |
|---|
-Details
| Has ligand of interest | Y |
|---|---|
| Has protein modification | N |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Value: 0.4 MDa / Experimental value: NO | ||||||||||||||||||||||||
| Source (natural) |
| ||||||||||||||||||||||||
| Source (recombinant) |
| ||||||||||||||||||||||||
| Buffer solution | pH: 7.5 | ||||||||||||||||||||||||
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES Details: The MCM recruitment reaction was reconstituted in vitro using purified H. sapiens proteins, a short DNA template and ATPgammaS. Four microlitres of the entire reaction was applied on a grid ...Details: The MCM recruitment reaction was reconstituted in vitro using purified H. sapiens proteins, a short DNA template and ATPgammaS. Four microlitres of the entire reaction was applied on a grid and incubated for 1 min at room temperature before blotting with filter paper for 5 s and plunge-freezing in liquid ethane. | ||||||||||||||||||||||||
| Specimen support | Grid material: GOLD / Grid mesh size: 300 divisions/in. / Grid type: UltrAuFoil R1.2/1.3 | ||||||||||||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK IV / Cryogen name: ETHANE / Humidity: 90 % / Chamber temperature: 295 K |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 130000 X / Nominal defocus max: 2500 nm / Nominal defocus min: 1000 nm |
| Specimen holder | Cryogen: NITROGEN |
| Image recording | Average exposure time: 9.4 sec. / Electron dose: 49.28 e/Å2 / Detector mode: COUNTING / Film or detector model: GATAN K2 SUMMIT (4k x 4k) / Num. of grids imaged: 1 / Num. of real images: 31569 |
| EM imaging optics | Energyfilter name: GIF Bioquantum |
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 4 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 182341 / Details: Local refinement in cryoSPARC / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building | PDB-ID: 7JPS Accession code: 7JPS Details: The PDB model was fit into the map and chains A to E were used as a starting model. Source name: PDB / Type: experimental model | ||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement | Cross valid method: NONE Stereochemistry target values: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 120.12 Å2 | ||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
|