- PDB-8pql: K48-linked ubiquitin chain formation with a cullin-RING E3 ligase... -
+
データを開く
IDまたはキーワード:
読み込み中...
-
基本情報
登録情報
データベース: PDB / ID: 8pql
タイトル
K48-linked ubiquitin chain formation with a cullin-RING E3 ligase and Cdc34: NEDD8-CUL2-RBX1-ELOB/C-FEM1C with trapped UBE2R2-donor UB-acceptor UB-SIL1 peptide
adenyl-nucleotide exchange factor activity / cotranslational protein targeting to membrane / negative regulation of beige fat cell differentiation / cullin-RING-type E3 NEDD8 transferase / NEDD8 transferase activity / cullin-RING ubiquitin ligase complex / Cul7-RING ubiquitin ligase complex / cellular response to chemical stress / Loss of Function of FBXW7 in Cancer and NOTCH1 Signaling / target-directed miRNA degradation ...adenyl-nucleotide exchange factor activity / cotranslational protein targeting to membrane / negative regulation of beige fat cell differentiation / cullin-RING-type E3 NEDD8 transferase / NEDD8 transferase activity / cullin-RING ubiquitin ligase complex / Cul7-RING ubiquitin ligase complex / cellular response to chemical stress / Loss of Function of FBXW7 in Cancer and NOTCH1 Signaling / target-directed miRNA degradation / elongin complex / positive regulation of protein autoubiquitination / RNA polymerase II transcription initiation surveillance / protein neddylation / NEDD8 ligase activity / protein K27-linked ubiquitination / negative regulation of response to oxidative stress / VCB complex / Cul5-RING ubiquitin ligase complex / ubiquitin-ubiquitin ligase activity / E2 ubiquitin-conjugating enzyme / ubiquitin-dependent protein catabolic process via the C-end degron rule pathway / SCF ubiquitin ligase complex / Cul2-RING ubiquitin ligase complex / Cul3-RING ubiquitin ligase complex / negative regulation of type I interferon production / SCF-dependent proteasomal ubiquitin-dependent protein catabolic process / Cul4A-RING E3 ubiquitin ligase complex / Cul4-RING E3 ubiquitin ligase complex / Prolactin receptor signaling / negative regulation of mitophagy / Cul4B-RING E3 ubiquitin ligase complex / ubiquitin ligase complex scaffold activity / ubiquitin conjugating enzyme activity / Pausing and recovery of Tat-mediated HIV elongation / Tat-mediated HIV elongation arrest and recovery / HIV elongation arrest and recovery / Pausing and recovery of HIV elongation / cullin family protein binding / protein monoubiquitination / ubiquitin ligase complex / Tat-mediated elongation of the HIV-1 transcript / Formation of HIV-1 elongation complex containing HIV-1 Tat / Formation of HIV elongation complex in the absence of HIV Tat / ubiquitin-like ligase-substrate adaptor activity / site of DNA damage / RNA Polymerase II Transcription Elongation / Formation of RNA Pol II elongation complex / signal transduction in response to DNA damage / Nuclear events stimulated by ALK signaling in cancer / protein K48-linked ubiquitination / transcription-coupled nucleotide-excision repair / negative regulation of insulin receptor signaling pathway / Maturation of protein E / Maturation of protein E / RNA Polymerase II Pre-transcription Events / regulation of cellular response to insulin stimulus / ER Quality Control Compartment (ERQC) / positive regulation of TORC1 signaling / Myoclonic epilepsy of Lafora / FLT3 signaling by CBL mutants / IRAK2 mediated activation of TAK1 complex / Prevention of phagosomal-lysosomal fusion / Alpha-protein kinase 1 signaling pathway / Glycogen synthesis / IRAK1 recruits IKK complex / IRAK1 recruits IKK complex upon TLR7/8 or 9 stimulation / Endosomal Sorting Complex Required For Transport (ESCRT) / Membrane binding and targetting of GAG proteins / Negative regulation of FLT3 / Regulation of TBK1, IKKε (IKBKE)-mediated activation of IRF3, IRF7 / PTK6 Regulates RTKs and Their Effectors AKT1 and DOK1 / Regulation of TBK1, IKKε-mediated activation of IRF3, IRF7 upon TLR3 ligation / IRAK2 mediated activation of TAK1 complex upon TLR7/8 or 9 stimulation / Constitutive Signaling by NOTCH1 HD Domain Mutants / NOTCH2 Activation and Transmission of Signal to the Nucleus / TICAM1,TRAF6-dependent induction of TAK1 complex / TICAM1-dependent activation of IRF3/IRF7 / post-translational protein modification / APC/C:Cdc20 mediated degradation of Cyclin B / intrinsic apoptotic signaling pathway / Regulation of FZD by ubiquitination / Downregulation of ERBB4 signaling / APC-Cdc20 mediated degradation of Nek2A / p75NTR recruits signalling complexes / InlA-mediated entry of Listeria monocytogenes into host cells / TRAF6 mediated IRF7 activation in TLR7/8 or 9 signaling / NF-kB is activated and signals survival / TRAF6-mediated induction of TAK1 complex within TLR4 complex / Regulation of pyruvate metabolism / Pexophagy / Regulation of innate immune responses to cytosolic DNA / NRIF signals cell death from the nucleus / Downregulation of ERBB2:ERBB3 signaling / Regulation of PTEN localization / VLDLR internalisation and degradation / Activated NOTCH1 Transmits Signal to the Nucleus / Synthesis of active ubiquitin: roles of E1 and E2 enzymes / T cell activation / Translesion synthesis by REV1 類似検索 - 分子機能
ジャーナル: Nat Struct Mol Biol / 年: 2024 タイトル: Mechanism of millisecond Lys48-linked poly-ubiquitin chain formation by cullin-RING ligases. 著者: Joanna Liwocha / Jerry Li / Nicholas Purser / Chutima Rattanasopa / Samuel Maiwald / David T Krist / Daniel C Scott / Barbara Steigenberger / J Rajan Prabu / Brenda A Schulman / Gary Kleiger / 要旨: E3 ubiquitin ligases, in collaboration with E2 ubiquitin-conjugating enzymes, modify proteins with poly-ubiquitin chains. Cullin-RING ligase (CRL) E3s use Cdc34/UBE2R-family E2s to build Lys48-linked ...E3 ubiquitin ligases, in collaboration with E2 ubiquitin-conjugating enzymes, modify proteins with poly-ubiquitin chains. Cullin-RING ligase (CRL) E3s use Cdc34/UBE2R-family E2s to build Lys48-linked poly-ubiquitin chains to control an enormous swath of eukaryotic biology. Yet the molecular mechanisms underlying this exceptional linkage specificity and millisecond kinetics of poly-ubiquitylation remain unclear. Here we obtain cryogenic-electron microscopy (cryo-EM) structures that provide pertinent insight into how such poly-ubiquitin chains are forged. The CRL RING domain not only activates the E2-bound ubiquitin but also shapes the conformation of a distinctive UBE2R2 loop, positioning both the ubiquitin to be transferred and the substrate-linked acceptor ubiquitin within the active site. The structures also reveal how the ubiquitin-like protein NEDD8 uniquely activates CRLs during chain formation. NEDD8 releases the RING domain from the CRL, but unlike previous CRL-E2 structures, does not contact UBE2R2. These findings suggest how poly-ubiquitylation may be accomplished by many E2s and E3s.
履歴
登録
2023年7月11日
登録サイト: PDBE / 処理サイト: PDBE
改定 1.0
2024年2月14日
Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: EM metadata / Data content type: EM metadata / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 1 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 2 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 3 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 4 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 5 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Image / Data content type: Image / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Primary map / Data content type: Primary map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 1 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 2 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 3 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 4 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 5 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Image / Data content type: Image / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Primary map / Data content type: Primary map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 1 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 2 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 3 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 4 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 5 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Image / Data content type: Image / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Primary map / Data content type: Primary map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 1 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 2 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 3 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 4 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 5 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Image / Data content type: Image / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Primary map / Data content type: Primary map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 1 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 2 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 3 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 4 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Additional map / Part number: 5 / Data content type: Additional map / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Image / Data content type: Image / Provider: repository / タイプ: Initial release
改定 1.0
2024年2月14日
Data content type: Primary map / Data content type: Primary map / Provider: repository / タイプ: Initial release
Data content type: EM metadata / Data content type: EM metadata / EM metadata / Group: Data processing / Experimental summary / Data content type: EM metadata / EM metadata / カテゴリ: em_admin / em_software / Data content type: EM metadata / EM metadata / Item: _em_admin.last_update / _em_software.name
E3ubiquitin-proteinligaseRBX1 / E3 ubiquitin-protein transferase RBX1 / Protein ZYP / RING finger protein 75 / RING-box protein 1 / ...E3 ubiquitin-protein transferase RBX1 / Protein ZYP / RING finger protein 75 / RING-box protein 1 / Rbx1 / Regulator of cullins 1 / ROC1
K48-linked ubiquitin chain formation with a cullin-RING E3 ligase and CDC34:NEDD8-CUL2-RBX1-ELOB/C-FEM1C with trapped UBE2R2-donor UB-acceptor UB-SIL1 peptide
COMPLEX
#1-#8
0
MULTIPLESOURCES
2
RING E3 ligase (RBX1) and Cullin-2 (CUL2)
COMPLEX
#4, #6
1
RECOMBINANT
3
CDC34, NEDD8, ELOB, ELOC, FEM1C with UBE2R2-donor UB-acceptor UB-SIL1 peptide and K48-linked ubiquitin chain
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト) データ登録者
データ登録者 ドイツ, 1件
ドイツ, 1件  引用
引用
 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード












 PDBj
PDBj



















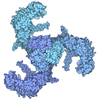






 集合体
集合体

 Trichoplusia ni (イラクサキンウワバ)
Trichoplusia ni (イラクサキンウワバ)

 分子量: 101.147 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C5H11NO
分子量: 101.147 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C5H11NO 分子量: 65.409 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Zn
分子量: 65.409 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Zn 試料調製
試料調製 電子顕微鏡撮影
電子顕微鏡撮影
 FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM
FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM 解析
解析