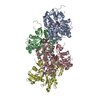
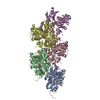
登録情報 データベース : PDB / ID : 8oidタイトル Cryo-EM structure of ADP-bound, filamentous beta-actin harboring the N111S mutation Actin, cytoplasmic 1, N-terminally processed キーワード / / / 機能・相同性 分子機能 ドメイン・相同性 構成要素
/ / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / 生物種 Homo sapiens (ヒト)手法 / / / 解像度 : 2.3 Å データ登録者 Oosterheert, W. / Blanc, F.E.C. / Roy, A. / Belyy, A. / Hofnagel, O. / Hummer, G. / Bieling, P. / Raunser, S. 資金援助 組織 認可番号 国 Max Planck Society Alexander von Humboldt Foundation European Research Council (ERC) 856118 European Union
ジャーナル : Nat Struct Mol Biol / 年 : 2023タイトル : Molecular mechanisms of inorganic-phosphate release from the core and barbed end of actin filaments.著者 : Wout Oosterheert / Florian E C Blanc / Ankit Roy / Alexander Belyy / Micaela Boiero Sanders / Oliver Hofnagel / Gerhard Hummer / Peter Bieling / Stefan Raunser / 要旨 : The release of inorganic phosphate (P) from actin filaments constitutes a key step in their regulated turnover, which is fundamental to many cellular functions. The mechanisms underlying P release ... The release of inorganic phosphate (P) from actin filaments constitutes a key step in their regulated turnover, which is fundamental to many cellular functions. The mechanisms underlying P release from the core and barbed end of actin filaments remain unclear. Here, using human and bovine actin isoforms, we combine cryo-EM with molecular-dynamics simulations and in vitro reconstitution to demonstrate how actin releases P through a 'molecular backdoor'. While constantly open at the barbed end, the backdoor is predominantly closed in filament-core subunits and opens only transiently through concerted amino acid rearrangements. This explains why P escapes rapidly from the filament end but slowly from internal subunits. In a nemaline-myopathy-associated actin variant, the backdoor is predominantly open in filament-core subunits, resulting in accelerated P release and filaments with drastically shortened ADP-P caps. Our results provide the molecular basis for P release from actin and exemplify how a disease-linked mutation distorts the nucleotide-state distribution and atomic structure of the filament. 履歴 登録 2023年3月22日 登録サイト / 処理サイト 改定 1.0 2023年8月9日 Provider / タイプ 改定 1.1 2023年8月16日 Group Data collection / Database references ... Data collection / Database references / Refinement description / Source and taxonomy / Structure summary カテゴリ chem_comp_atom / chem_comp_bond ... chem_comp_atom / chem_comp_bond / citation / entity / entity_src_gen / struct_ncs_dom_lim / struct_ref / struct_ref_seq / struct_ref_seq_dif Item _citation.country / _citation.journal_abbrev ... _citation.country / _citation.journal_abbrev / _citation.journal_id_CSD / _citation.journal_id_ISSN / _citation.year / _entity.pdbx_description / _entity_src_gen.pdbx_gene_src_gene / _struct_ncs_dom_lim.beg_auth_asym_id / _struct_ncs_dom_lim.beg_auth_comp_id / _struct_ncs_dom_lim.beg_auth_seq_id / _struct_ncs_dom_lim.beg_label_asym_id / _struct_ncs_dom_lim.beg_label_seq_id / _struct_ncs_dom_lim.end_auth_asym_id / _struct_ncs_dom_lim.end_auth_comp_id / _struct_ncs_dom_lim.end_auth_seq_id / _struct_ncs_dom_lim.end_label_asym_id / _struct_ncs_dom_lim.end_label_seq_id / _struct_ref.db_code / _struct_ref.pdbx_db_accession / _struct_ref_seq.pdbx_db_accession / _struct_ref_seq_dif.pdbx_seq_db_accession_code 改定 1.2 2023年9月27日 Group / Refinement descriptionカテゴリ / citation_author / em_3d_fitting_listItem / _em_3d_fitting_list.initial_refinement_model_id改定 1.3 2023年10月4日 Group / カテゴリ / citation_authorItem / _citation.title / _citation_author.identifier_ORCID改定 1.4 2023年11月22日 Group / カテゴリ / citation_authorItem _citation.journal_volume / _citation.page_first ... _citation.journal_volume / _citation.page_first / _citation.page_last / _citation_author.identifier_ORCID
すべて表示 表示を減らす
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト) データ登録者
データ登録者 ドイツ, European Union, 3件
ドイツ, European Union, 3件  引用
引用 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード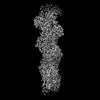


 PDBj
PDBj
















 集合体
集合体