 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-8i7o: In situ structure of axonemal doublet microtubules in mouse sperm... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8i7o | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | In situ structure of axonemal doublet microtubules in mouse sperm with 16-nm repeat | |||||||||||||||||||||
 Components Components |
| |||||||||||||||||||||
 Keywords Keywords | STRUCTURAL PROTEIN / microtubules / axoneme / sperm / filament | |||||||||||||||||||||
| Function / homology |  Function and homology information Function and homology informationouter acrosomal membrane / regulation of brood size / establishment of left/right asymmetry / 9+0 motile cilium / axonemal microtubule doublet inner sheath / axonemal B tubule inner sheath / axonemal A tubule inner sheath / Microtubule-dependent trafficking of connexons from Golgi to the plasma membrane / Cilium Assembly / Sealing of the nuclear envelope (NE) by ESCRT-III ...outer acrosomal membrane / regulation of brood size / establishment of left/right asymmetry / 9+0 motile cilium / axonemal microtubule doublet inner sheath / axonemal B tubule inner sheath / axonemal A tubule inner sheath / Microtubule-dependent trafficking of connexons from Golgi to the plasma membrane / Cilium Assembly / Sealing of the nuclear envelope (NE) by ESCRT-III / protein polyglutamylation / Intraflagellar transport / Carboxyterminal post-translational modifications of tubulin / positive regulation of feeding behavior / COPI-independent Golgi-to-ER retrograde traffic / MAP kinase tyrosine/serine/threonine phosphatase activity / inner dynein arm assembly / cilium-dependent cell motility / regulation of cilium beat frequency involved in ciliary motility / HSP90 chaperone cycle for steroid hormone receptors (SHR) in the presence of ligand / COPI-mediated anterograde transport / cilium movement involved in cell motility / 9+2 motile cilium / Kinesins / acrosomal membrane / axoneme assembly / PKR-mediated signaling / Aggrephagy / RHO GTPases activate IQGAPs / microtubule sliding / Mitotic Prometaphase / EML4 and NUDC in mitotic spindle formation / Resolution of Sister Chromatid Cohesion / Recycling pathway of L1 / The role of GTSE1 in G2/M progression after G2 checkpoint / COPI-dependent Golgi-to-ER retrograde traffic / axonemal microtubule / Hedgehog 'off' state / RHO GTPases Activate Formins / Loss of Nlp from mitotic centrosomes / Recruitment of mitotic centrosome proteins and complexes / Loss of proteins required for interphase microtubule organization from the centrosome / Anchoring of the basal body to the plasma membrane / Separation of Sister Chromatids / cilium organization / Recruitment of NuMA to mitotic centrosomes / AURKA Activation by TPX2 / manchette / Regulation of PLK1 Activity at G2/M Transition / MHC class II antigen presentation / flagellated sperm motility / motile cilium / extrinsic component of membrane / protein targeting to membrane / positive regulation of cell motility / : / tubulin complex / protein-serine/threonine phosphatase / regulation of neuron projection development / ciliary base / protein serine/threonine phosphatase activity / cerebral cortex cell migration / phosphatase activity / mitotic cytokinesis / microtubule organizing center / cilium assembly / cellular response to unfolded protein / spermatid development / regulation of cell division / single fertilization / axoneme / sperm flagellum / alpha-tubulin binding / microtubule-based process / intercellular bridge / beta-tubulin binding / cytoplasmic microtubule / phosphoprotein phosphatase activity / protein-tyrosine-phosphatase / heat shock protein binding / Neutrophil degranulation / protein tyrosine phosphatase activity / Hsp70 protein binding / acrosomal vesicle / mitotic spindle organization / cellular response to leukemia inhibitory factor / centriole / Hsp90 protein binding / SH3 domain binding / mitochondrial intermembrane space / structural constituent of cytoskeleton / G protein-coupled receptor binding / microtubule cytoskeleton organization / intracellular calcium ion homeostasis / spindle pole / calcium-dependent protein binding / mitotic spindle / intracellular protein localization / myelin sheath / mitotic cell cycle Similarity search - Function | |||||||||||||||||||||
| Biological species |  | |||||||||||||||||||||
| Method | ELECTRON MICROSCOPY / subtomogram averaging / cryo EM / Resolution: 4.5 Å | |||||||||||||||||||||
 Authors Authors | Zhu, Y. / Yin, G.L. / Tai, L.H. / Sun, F. | |||||||||||||||||||||
| Funding support |  China, 1items China, 1items
| |||||||||||||||||||||
 Citation Citation | Journal: Cell Discov / Year: 2023 Title: In-cell structural insight into the stability of sperm microtubule doublet. Authors: Linhua Tai / Guoliang Yin / Xiaojun Huang / Fei Sun / Yun Zhu / Abstract: The propulsion for mammalian sperm swimming is generated by flagella beating. Microtubule doublets (DMTs) along with microtubule inner proteins (MIPs) are essential structural blocks of flagella. ...The propulsion for mammalian sperm swimming is generated by flagella beating. Microtubule doublets (DMTs) along with microtubule inner proteins (MIPs) are essential structural blocks of flagella. However, the intricate molecular architecture of intact sperm DMT remains elusive. Here, by in situ cryo-electron tomography, we solved the in-cell structure of mouse sperm DMT at 4.5-7.5 Å resolutions, and built its model with 36 kinds of MIPs in 48 nm periodicity. We identified multiple copies of Tektin5 that reinforce Tektin bundle, and multiple MIPs with different periodicities that anchor the Tektin bundle to tubulin wall. This architecture contributes to a superior stability of A-tubule than B-tubule of DMT, which was revealed by structural comparison of DMTs from the intact and deformed axonemes. Our work provides an overall molecular picture of intact sperm DMT in 48 nm periodicity that is essential to understand the molecular mechanism of sperm motility as well as the related ciliopathies. | |||||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8i7o.cif.gz | 11.4 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8i7o.ent.gz | Display | PDB format | |
| PDBx/mmJSON format | 8i7o.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/i7/8i7oftp://data.pdbj.org/pub/pdb/validation_reports/i7/8i7o | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  35229MC  8i7rC C: citing same article ( M: map data used to model this data |
|---|---|
| Similar structure data |















-Links
 PDBj
PDBj




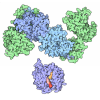











- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-Protein , 19 types, 179 molecules A2A3AEAGAIBEBGBICGCIDEDGEEEGFEFGGEGGGIHEHGHIIEIGIIJEJGKEKGKI...
| #1: Protein | Mass: 48713.035 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #2: Protein | Mass: 48689.090 Da / Num. of mol.: 61 / Source method: isolated from a natural source / Source: (natural) #3: Protein | Mass: 47897.918 Da / Num. of mol.: 62 / Source method: isolated from a natural source / Source: (natural) #4: Protein | Mass: 50385.066 Da / Num. of mol.: 4 / Source method: isolated from a natural source / Source: (natural) #5: Protein | Mass: 56754.777 Da / Num. of mol.: 7 / Source method: isolated from a natural source / Source: (natural) #6: Protein | Mass: 52121.449 Da / Num. of mol.: 6 / Source method: isolated from a natural source / Source: (natural) #7: Protein | Mass: 23987.262 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #8: Protein | Mass: 62817.535 Da / Num. of mol.: 8 / Source method: isolated from a natural source / Source: (natural) #11: Protein | Mass: 36659.566 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #12: Protein | Mass: 23097.957 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #13: Protein | Mass: 21527.574 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) References: UniProt: Q9D9D8, protein-serine/threonine phosphatase, protein-tyrosine-phosphatase #14: Protein | Mass: 57406.484 Da / Num. of mol.: 3 / Source method: isolated from a natural source / Source: (natural) #15: Protein | Mass: 29587.521 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #16: Protein | Mass: 16305.608 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #18: Protein | Mass: 20575.123 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #20: Protein | Mass: 176028.312 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #21: Protein | | Mass: 32786.020 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) #23: Protein | Mass: 27763.268 Da / Num. of mol.: 3 / Source method: isolated from a natural source / Source: (natural) #24: Protein | Mass: 19512.373 Da / Num. of mol.: 6 / Source method: isolated from a natural source / Source: (natural) |
|---|
-EF-hand domain-containing ... , 2 types, 3 molecules G1G2G5
| #9: Protein | Mass: 75235.422 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #10: Protein | | Mass: 87758.023 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|
-Cilia- and flagella-associated protein ... , 3 types, 7 molecules N2N3P1P2XCXDXE
| #17: Protein | Mass: 18960.092 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #19: Protein | Mass: 68322.164 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #22: Protein | Mass: 22781.389 Da / Num. of mol.: 3 / Source method: isolated from a natural source / Source: (natural) |
|---|
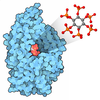
-Non-polymers , 1 types, 123 molecules 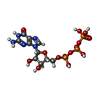
| #25: Chemical | ChemComp-GTP /  Mass: 523.180 Da / Num. of mol.: 123 / Source method: obtained synthetically / Formula: C10H16N5O14P3 / Comment: GTP, energy-carrying molecule*YM Mass: 523.180 Da / Num. of mol.: 123 / Source method: obtained synthetically / Formula: C10H16N5O14P3 / Comment: GTP, energy-carrying molecule*YM |
|---|
-Details
| Has ligand of interest | N |
|---|---|
| Has protein modification | Y |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: CELL / 3D reconstruction method: subtomogram averaging |
- Sample preparation
Sample preparation
| Component | Name: mouse sperm / Type: CELL / Entity ID: #1-#24 / Source: NATURAL |
|---|---|
| Source (natural) | Organism: |
| Buffer solution | pH: 7 |
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: SPOT SCAN FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: SPOT SCAN |
| Electron lens | Mode: BRIGHT FIELD / Nominal defocus max: 5000 nm / Nominal defocus min: 1000 nm / Cs: 2.7 mm |
| Image recording | Electron dose: 3 e/Å2 / Avg electron dose per subtomogram: 117 e/Å2 / Detector mode: SUPER-RESOLUTION / Film or detector model: GATAN K2 QUANTUM (4k x 4k) |
- Processing
Processing
| Software | Name: UCSF ChimeraX / Version: 1.6/v9 / Classification: model building / URL: https://www.rbvi.ucsf.edu/chimerax/ / Os: Windows / Type: package |
|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION |
| Symmetry | Point symmetry: C1 (asymmetric) |
| 3D reconstruction | Resolution: 4.5 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 37018 / Symmetry type: POINT |
| EM volume selection | Num. of tomograms: 689 / Num. of volumes extracted: 37018 |
