 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 7tqy | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | CaKip3[2-482] - ADP-AlFx in complex with a microtubule | ||||||||||||
 Components Components |
| ||||||||||||
 Keywords Keywords | MOTOR PROTEIN / Kip3 / kinesin / motility / microtubule / tubulin | ||||||||||||
| Function / homology |  Function and homology information Function and homology informationplus-end specific microtubule depolymerization / tubulin-dependent ATPase activity / regulation of mitotic spindle elongation / mitotic spindle astral microtubule / meiotic sister chromatid segregation / nuclear microtubule / mitotic spindle midzone / nuclear migration along microtubule / microtubule plus-end / microtubule nucleation ...plus-end specific microtubule depolymerization / tubulin-dependent ATPase activity / regulation of mitotic spindle elongation / mitotic spindle astral microtubule / meiotic sister chromatid segregation / nuclear microtubule / mitotic spindle midzone / nuclear migration along microtubule / microtubule plus-end / microtubule nucleation / mitotic spindle disassembly / plus-end-directed microtubule motor activity / Microtubule-dependent trafficking of connexons from Golgi to the plasma membrane / Resolution of Sister Chromatid Cohesion / Hedgehog 'off' state / Cilium Assembly / Intraflagellar transport / COPI-dependent Golgi-to-ER retrograde traffic / Mitotic Prometaphase / Carboxyterminal post-translational modifications of tubulin / RHOH GTPase cycle / EML4 and NUDC in mitotic spindle formation / Sealing of the nuclear envelope (NE) by ESCRT-III / Kinesins / PKR-mediated signaling / Separation of Sister Chromatids / The role of GTSE1 in G2/M progression after G2 checkpoint / Aggrephagy / RHO GTPases activate IQGAPs / RHO GTPases Activate Formins / kinesin complex / microtubule depolymerization / HSP90 chaperone cycle for steroid hormone receptors (SHR) in the presence of ligand / MHC class II antigen presentation / Recruitment of NuMA to mitotic centrosomes / COPI-mediated anterograde transport / microtubule-based movement / negative regulation of microtubule polymerization / establishment of mitotic spindle orientation / mitotic sister chromatid segregation / mitotic spindle assembly / microtubule-based process / mitotic spindle organization / structural constituent of cytoskeleton / microtubule cytoskeleton organization / mitotic cell cycle / microtubule cytoskeleton / microtubule binding / Hydrolases; Acting on acid anhydrides; Acting on GTP to facilitate cellular and subcellular movement / microtubule / GTPase activity / GTP binding / ATP hydrolysis activity / ATP binding / metal ion binding / nucleus / cytoplasm Similarity search - Function | ||||||||||||
| Biological species |  Candida albicans (yeast) Candida albicans (yeast) | ||||||||||||
| Method | ELECTRON MICROSCOPY / helical reconstruction / cryo EM / Resolution: 2.6 Å | ||||||||||||
 Authors Authors | Benoit, M.P.M.H. / Asenjo, A.B. / Hunter, B. / Allingham, J. / Sosa, H. | ||||||||||||
| Funding support |  United States, United States,  Canada, 3items Canada, 3items
| ||||||||||||
 Citation Citation | Journal: Nat Commun / Year: 2022 Title: Kinesin-8-specific loop-2 controls the dual activities of the motor domain according to tubulin protofilament shape. Authors: Byron Hunter / Matthieu P M H Benoit / Ana B Asenjo / Caitlin Doubleday / Daria Trofimova / Corey Frazer / Irsa Shoukat / Hernando Sosa / John S Allingham / Abstract: Kinesin-8s are dual-activity motor proteins that can move processively on microtubules and depolymerize microtubule plus-ends, but their mechanism of combining these distinct activities remains ...Kinesin-8s are dual-activity motor proteins that can move processively on microtubules and depolymerize microtubule plus-ends, but their mechanism of combining these distinct activities remains unclear. We addressed this by obtaining cryo-EM structures (2.6-3.9 Å) of Candida albicans Kip3 in different catalytic states on the microtubule lattice and on a curved microtubule end mimic. We also determined a crystal structure of microtubule-unbound CaKip3-ADP (2.0 Å) and analyzed the biochemical activity of CaKip3 and kinesin-1 mutants. These data reveal that the microtubule depolymerization activity of kinesin-8 originates from conformational changes of its motor core that are amplified by dynamic contacts between its extended loop-2 and tubulin. On curved microtubule ends, loop-1 inserts into preceding motor domains, forming head-to-tail arrays of kinesin-8s that complement loop-2 contacts with curved tubulin and assist depolymerization. On straight tubulin protofilaments in the microtubule lattice, loop-2-tubulin contacts inhibit conformational changes in the motor core, but in the ADP-Pi state these contacts are relaxed, allowing neck-linker docking for motility. We propose that these tubulin shape-induced alternations between pro-microtubule-depolymerization and pro-motility kinesin states, regulated by loop-2, are the key to the dual activity of kinesin-8 motors. | ||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 7tqy.cif.gz | 261.1 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb7tqy.ent.gz | 201.4 KB | Display | PDB format |
| PDBx/mmJSON format | 7tqy.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Summary document | 7tqy_validation.pdf.gz | 1.8 MB | Display | wwPDB validaton report |
|---|---|---|---|---|
| Full document | 7tqy_full_validation.pdf.gz | 1.8 MB | Display | |
| Data in XML | 7tqy_validation.xml.gz | 53.9 KB | Display | |
| Data in CIF | 7tqy_validation.cif.gz | 79.3 KB | Display | |
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/tq/7tqyftp://data.pdbj.org/pub/pdb/validation_reports/tq/7tqy | HTTPS FTP |
-Related structure data
| Related structure data |  26075MC  7lffC  7tqxC  7tqzC  7tr0C  7tr1C  7tr2C  7tr3C M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |






-Links
 PDBj
PDBj






- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-Protein , 3 types, 3 molecules KAB
| #1: Protein | Mass: 54869.469 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Candida albicans (yeast) / Gene: KIP3, orf19.7353, CAALFM_C305720CA / Production host:  |
|---|---|
| #2: Protein | Mass: 50204.445 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #3: Protein | Mass: 49999.887 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
-Non-polymers , 6 types, 7 molecules 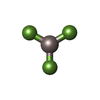


| #4: Chemical | ChemComp-AF3 /  Mass: 83.977 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: AlF3 Mass: 83.977 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: AlF3 | ||||||
|---|---|---|---|---|---|---|---|
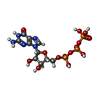
| #5: Chemical | ChemComp-ADP /  Mass: 427.201 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Comment: ADP, energy-carrying molecule*YM Mass: 427.201 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Comment: ADP, energy-carrying molecule*YM | ||||||
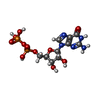
| #6: Chemical |  Mass: 24.305 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Mg#7: Chemical | ChemComp-GTP / |  Mass: 523.180 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H16N5O14P3 / Comment: GTP, energy-carrying molecule*YM Mass: 523.180 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H16N5O14P3 / Comment: GTP, energy-carrying molecule*YM#8: Chemical | ChemComp-GDP / |  Type: RNA linking / Mass: 443.201 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H15N5O11P2 / Comment: GDP, energy-carrying molecule*YM Type: RNA linking / Mass: 443.201 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H15N5O11P2 / Comment: GDP, energy-carrying molecule*YM#9: Chemical | ChemComp-TA1 / |  Mass: 853.906 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C47H51NO14 / Comment: medication, chemotherapy*YM Mass: 853.906 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C47H51NO14 / Comment: medication, chemotherapy*YM |
-Details
| Has ligand of interest | N |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: FILAMENT / 3D reconstruction method: helical reconstruction |
- Sample preparation
Sample preparation
| Component |
| ||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Experimental value: NO | ||||||||||||||||||||||||||||||||||||||||
| Source (natural) |
| ||||||||||||||||||||||||||||||||||||||||
| Source (recombinant) | Organism: | ||||||||||||||||||||||||||||||||||||||||
| Buffer solution | pH: 6.8 | ||||||||||||||||||||||||||||||||||||||||
| Buffer component |
| ||||||||||||||||||||||||||||||||||||||||
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||||||||||||||||||||||
| Specimen support | Grid material: GOLD / Grid type: UltrAuFoil R2/2 | ||||||||||||||||||||||||||||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK IV / Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal defocus max: 1500 nm / Nominal defocus min: 750 nm / Alignment procedure: COMA FREE |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Electron dose: 63.28 e/Å2 / Detector mode: COUNTING / Film or detector model: GATAN K2 SUMMIT (4k x 4k) |
| Image scans | Movie frames/image: 40 / Used frames/image: 1-40 |
- Processing
Processing
| EM software |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Helical symmerty | Angular rotation/subunit: 168.069 ° / Axial rise/subunit: 5.44 Å / Axial symmetry: C1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Particle selection | Details: manual picking of filaments | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 2.6 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 255526 Details: 2 half datasets containing one distinct half of each filament were refined independently. Number of asymmetric units used is reported in "number of segments used", due to the local processing strategy employed. Symmetry type: HELICAL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building | Protocol: FLEXIBLE FIT / Space: REAL |
