 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 7nfe | ||||||
|---|---|---|---|---|---|---|---|
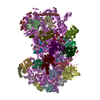
| Title | Cryo-EM structure of NHEJ super-complex (monomer) | ||||||
 Components Components |
| ||||||
 Keywords Keywords | DNA BINDING PROTEIN / NHEJ / DNA-PKcs / Ku70/80 / XLF / XRCC4 / DNA-LigaseIV | ||||||
| Function / homology |  Function and homology information Function and homology informationFHA domain binding / positive regulation of chromosome organization / DN2 thymocyte differentiation / positive regulation of ligase activity / DNA ligase IV complex / positive regulation of platelet formation / DNA ligase activity / DNA double-strand break attachment to nuclear envelope / Ku70:Ku80 complex / T cell receptor V(D)J recombination ...FHA domain binding / positive regulation of chromosome organization / DN2 thymocyte differentiation / positive regulation of ligase activity / DNA ligase IV complex / positive regulation of platelet formation / DNA ligase activity / DNA double-strand break attachment to nuclear envelope / Ku70:Ku80 complex / T cell receptor V(D)J recombination / DNA ligase (ATP) / pro-B cell differentiation / negative regulation of t-circle formation / DNA end binding / DNA-dependent protein kinase activity / DNA ligase (ATP) activity / small-subunit processome assembly / positive regulation of lymphocyte differentiation / histone H2AXS139 kinase activity / DNA-dependent protein kinase complex / DNA-dependent protein kinase-DNA ligase 4 complex / immunoglobulin V(D)J recombination / nonhomologous end joining complex / nucleotide-excision repair, DNA gap filling / single strand break repair / immature B cell differentiation / V(D)J recombination / regulation of smooth muscle cell proliferation / cellular response to X-ray / regulation of epithelial cell proliferation / double-strand break repair via classical nonhomologous end joining / isotype switching / double-strand break repair via alternative nonhomologous end joining / nuclear telomere cap complex / Cytosolic sensors of pathogen-associated DNA / protein localization to site of double-strand break / telomere capping / IRF3-mediated induction of type I IFN / regulation of hematopoietic stem cell differentiation / recombinational repair / regulation of telomere maintenance / protein localization to chromosome, telomeric region / U3 snoRNA binding / peptidyl-threonine phosphorylation / positive regulation of neurogenesis / maturation of 5.8S rRNA / T cell lineage commitment / cellular response to lithium ion / cellular hyperosmotic salinity response / DNA biosynthetic process / positive regulation of double-strand break repair via nonhomologous end joining / 2-LTR circle formation / B cell lineage commitment / negative regulation of cGAS/STING signaling pathway / AMP binding / response to ionizing radiation / hematopoietic stem cell proliferation / telomeric repeat DNA binding / ligase activity / negative regulation of protein phosphorylation / DNA 3'-5' helicase / T cell differentiation / somatic stem cell population maintenance / 5'-deoxyribose-5-phosphate lyase activity / chromosome organization / hematopoietic stem cell differentiation / response to X-ray / ectopic germ cell programmed cell death / somitogenesis / ATP-dependent activity, acting on DNA / telomere maintenance via telomerase / SUMOylation of DNA damage response and repair proteins / site of DNA damage / condensed chromosome / mitotic G1 DNA damage checkpoint signaling / DNA polymerase binding / activation of innate immune response / neurogenesis / positive regulation of erythrocyte differentiation / B cell differentiation / telomere maintenance / cyclin binding / stem cell proliferation / DNA-(apurinic or apyrimidinic site) lyase / peptidyl-serine phosphorylation / protein modification process / DNA helicase activity / cellular response to leukemia inhibitory factor / cellular response to ionizing radiation / response to gamma radiation / central nervous system development / positive regulation of translation / Nonhomologous End-Joining (NHEJ) / small-subunit processome / cellular response to gamma radiation / protein-DNA complex / regulation of circadian rhythm / brain development / base-excision repair / establishment of integrated proviral latency Similarity search - Function | ||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 4.29 Å | ||||||
 Authors Authors | Chaplin, A.K. / Hardwick, S.W. / Kefala Stavridi, A. / Chirgadze, D.Y. / Blundell, T.L. | ||||||
| Funding support |  United Kingdom, 1items United Kingdom, 1items
| ||||||
 Citation Citation | Journal: Mol Cell / Year: 2021 Title: Cryo-EM of NHEJ supercomplexes provides insights into DNA repair. Authors: Amanda K Chaplin / Steven W Hardwick / Antonia Kefala Stavridi / Christopher J Buehl / Noah J Goff / Virginie Ropars / Shikang Liang / Taiana Maia De Oliveira / Dimitri Y Chirgadze / ...Authors: Amanda K Chaplin / Steven W Hardwick / Antonia Kefala Stavridi / Christopher J Buehl / Noah J Goff / Virginie Ropars / Shikang Liang / Taiana Maia De Oliveira / Dimitri Y Chirgadze / Katheryn Meek / Jean-Baptiste Charbonnier / Tom L Blundell /   Abstract: Non-homologous end joining (NHEJ) is one of two critical mechanisms utilized in humans to repair DNA double-strand breaks (DSBs). Unrepaired or incorrect repair of DSBs can lead to apoptosis or ...Non-homologous end joining (NHEJ) is one of two critical mechanisms utilized in humans to repair DNA double-strand breaks (DSBs). Unrepaired or incorrect repair of DSBs can lead to apoptosis or cancer. NHEJ involves several proteins, including the Ku70/80 heterodimer, DNA-dependent protein kinase catalytic subunit (DNA-PKcs), X-ray cross-complementing protein 4 (XRCC4), XRCC4-like factor (XLF), and ligase IV. These core proteins bind DSBs and ligate the damaged DNA ends. However, details of the structural assembly of these proteins remain unclear. Here, we present cryo-EM structures of NHEJ supercomplexes that are composed of these core proteins and DNA, revealing the detailed structural architecture of this assembly. We describe monomeric and dimeric forms of this supercomplex and also propose the existence of alternate dimeric forms of long-range synaptic complexes. Finally, we show that mutational disruption of several structural features within these NHEJ complexes negatively affects DNA repair. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 7nfe.cif.gz | 1 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb7nfe.ent.gz | 808.7 KB | Display | PDB format |
| PDBx/mmJSON format | 7nfe.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/nf/7nfeftp://data.pdbj.org/pub/pdb/validation_reports/nf/7nfe | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  12301MC  7nfcC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |


-Links
 PDBj
PDBj












































- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-Protein , 4 types, 6 molecules AFGHIJ
| #1: Protein | Mass: 472056.281 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / Cell line: HELAReferences: UniProt: P78527, non-specific serine/threonine protein kinase | ||||
|---|---|---|---|---|---|
| #4: Protein | Mass: 33372.234 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: NHEJ1, XLF / Production host:  #5: Protein | Mass: 38337.703 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: XRCC4 / Production host: #6: Protein | | Mass: 104124.953 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: LIG4 / Production host: |
-X-ray repair cross-complementing protein ... , 2 types, 2 molecules BC
| #2: Protein | Mass: 69945.039 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: XRCC6, G22P1Production host: Insect cell expression vector pTIE1 (others) References: UniProt: P12956, Hydrolases; Acting on acid anhydrides; Acting on acid anhydrides to facilitate cellular and subcellular movement, Lyases; Carbon-oxygen lyases; Other carbon-oxygen lyases |
|---|---|
| #3: Protein | Mass: 82812.438 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: XRCC5, G22P2Production host: Insect cell expression vector pTIE1 (others) References: UniProt: P13010, Hydrolases; Acting on acid anhydrides; Acting on acid anhydrides to facilitate cellular and subcellular movement |
-DNA chain , 2 types, 2 molecules DE
| #7: DNA chain | Mass: 7367.843 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) Homo sapiens (human) |
|---|---|
| #8: DNA chain | Mass: 7402.821 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) Homo sapiens (human) |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component |
| ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Value: 0.8 MDa / Experimental value: NO | ||||||||||||||||||||||||||||||||||||
| Source (natural) |
| ||||||||||||||||||||||||||||||||||||
| Source (recombinant) |
| ||||||||||||||||||||||||||||||||||||
| Buffer solution | pH: 7.4 | ||||||||||||||||||||||||||||||||||||
| Specimen | Conc.: 3 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||||||||||||||||||
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 130000 X / Cs: 2.7 mm / C2 aperture diameter: 50 µm |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Average exposure time: 1.3 sec. / Electron dose: 46.8 e/Å2 / Film or detector model: GATAN K3 BIOQUANTUM (6k x 4k) |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM software |
| ||||||||||||||||||||||||
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 749185 | ||||||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | ||||||||||||||||||||||||
| 3D reconstruction | Resolution: 4.29 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 45943 / Symmetry type: POINT | ||||||||||||||||||||||||
| Refinement | Cross valid method: NONE Stereochemistry target values: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||
| Displacement parameters | Biso mean: 358.24 Å2 | ||||||||||||||||||||||||
| Refine LS restraints |
|
