 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 6xu6 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Drosophila melanogaster Testis 80S ribosome | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | RIBOSOME / Specialised ribosome / Drosophila melanogaster / Testis | |||||||||
| Function / homology |  Function and homology information Function and homology informationAPC/C:Cdc20 mediated degradation of Cyclin B / SCF-beta-TrCP mediated degradation of Emi1 / SRP-dependent cotranslational protein targeting to membrane / Activation of the IkappaB kinase complex, KEY:IRD5 dimer:KEY / NF-kB is activated and signals survival / Downregulation of TGF-beta receptor signaling / SMAD2/SMAD3:SMAD4 heterotrimer regulates transcription / Regulation of innate immune responses to cytosolic DNA / Major pathway of rRNA processing in the nucleolus and cytosol / Regulation of TP53 Degradation ...APC/C:Cdc20 mediated degradation of Cyclin B / SCF-beta-TrCP mediated degradation of Emi1 / SRP-dependent cotranslational protein targeting to membrane / Activation of the IkappaB kinase complex, KEY:IRD5 dimer:KEY / NF-kB is activated and signals survival / Downregulation of TGF-beta receptor signaling / SMAD2/SMAD3:SMAD4 heterotrimer regulates transcription / Regulation of innate immune responses to cytosolic DNA / Major pathway of rRNA processing in the nucleolus and cytosol / Regulation of TP53 Degradation / Cyclin D associated events in G1 / Formation of a pool of free 40S subunits / Formation of the ternary complex, and subsequently, the 43S complex / GTP hydrolysis and joining of the 60S ribosomal subunit / PTK6 Regulates RTKs and Their Effectors AKT1 and DOK1 / Protein methylation / ER Quality Control Compartment (ERQC) / Iron uptake and transport / RAS processing / Pexophagy / Regulation of BACH1 activity / Nonsense Mediated Decay (NMD) independent of the Exon Junction Complex (EJC) / : / Regulation of pyruvate metabolism / Translesion synthesis by REV1 / Downregulation of SMAD2/3:SMAD4 transcriptional activity / N-glycan trimming in the ER and Calnexin/Calreticulin cycle / Translesion synthesis by POLK / Translesion synthesis by POLI / Ovarian tumor domain proteases / Formation of Incision Complex in GG-NER / Regulation of expression of SLITs and ROBOs / Translesion Synthesis by POLH / Oxygen-dependent proline hydroxylation of Hypoxia-inducible Factor Alpha / Autodegradation of Cdh1 by Cdh1:APC/C / APC/C:Cdc20 mediated degradation of Securin / APC/C:Cdh1 mediated degradation of Cdc20 and other APC/C:Cdh1 targeted proteins in late mitosis/early G1 / APC-Cdc20 mediated degradation of Nek2A / SCF(Skp2)-mediated degradation of p27/p21 / Degradation of beta-catenin by the destruction complex / Ubiquitination and degradation of phosphorylated ARM / Separation of Sister Chromatids / Degradation of AXIN / Degradation of DVL / Regulation of TNFR1 signaling / Dectin-1 mediated noncanonical NF-kB signaling / Regulation of RAS by GAPs / Negative regulation of MAPK pathway / NIK-->noncanonical NF-kB signaling / Ubiquitin-Mediated Degradation of Phosphorylated Cdc25A / Ubiquitin-dependent degradation of Cyclin D / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis / Regulation of RUNX2 expression and activity / Regulation of RUNX3 expression and activity / Peroxisomal protein import / GSK3B and BTRC:CUL1-mediated-degradation of NFE2L2 / Recognition of DNA damage by PCNA-containing replication complex / Downregulation of ERBB2:ERBB3 signaling / Activated NOTCH1 Transmits Signal to the Nucleus / Nuclear CI is degraded / ABC-family protein mediated transport / TAK1-dependent IKK and NF-kappa-B activation / activated TAK1 mediates p38 MAPK activation / PINK1-PRKN Mediated Mitophagy / Josephin domain DUBs / Dual Incision in GG-NER / Downregulation of ERBB2 signaling / VLDLR internalisation and degradation / Endosomal Sorting Complex Required For Transport (ESCRT) / IRAK1 recruits IKK complex / IRAK2 mediated activation of TAK1 complex / TRAF6-mediated induction of TAK1 complex within TLR4 complex / IRAK1 recruits IKK complex upon TLR7/8 or 9 stimulation / IRAK2 mediated activation of TAK1 complex upon TLR7/8 or 9 stimulation / KEAP1-NFE2L2 pathway / Cdc20:Phospho-APC/C mediated degradation of Cyclin A / Ubiquitination and proteolysis of phosphorylated CI / JNK (c-Jun kinases) phosphorylation and activation mediated by activated human TAK1 / RUNX1 regulates transcription of genes involved in differentiation of HSCs / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / Interleukin-1 signaling / Degradation of CRY and PER proteins / pupariation / Downregulation of ERBB4 signaling / Spry regulation of FGF signaling / Degradation of GLI1 by the proteasome / GLI3 is processed to GLI3R by the proteasome / Negative regulation of FGFR1 signaling / Negative regulation of FGFR2 signaling / Negative regulation of FGFR3 signaling / Negative regulation of FGFR4 signaling / Gap-filling DNA repair synthesis and ligation in TC-NER / Synthesis of active ubiquitin: roles of E1 and E2 enzymes / Regulation of signaling by CBL / Protein hydroxylation / TGF-beta receptor signaling in EMT (epithelial to mesenchymal transition) / Formation of TC-NER Pre-Incision Complex / Deactivation of the beta-catenin transactivating complex / Hedgehog ligand biogenesis / Regulation of necroptotic cell death Similarity search - Function | |||||||||
| Biological species |  | |||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.5 Å | |||||||||
 Authors Authors | Hopes, T. / Agapiou, M. / Norris, K. / McCarthy, C.G.P. / OConnell, M.J. / Fontana, J. / Aspden, J.L. | |||||||||
| Funding support |  United Kingdom, 2items United Kingdom, 2items
| |||||||||
 Citation Citation | Journal: Nucleic Acids Res / Year: 2022 Title: Ribosome heterogeneity in Drosophila melanogaster gonads through paralog-switching. Authors: Tayah Hopes / Karl Norris / Michaela Agapiou / Charley G P McCarthy / Philip A Lewis / Mary J O'Connell / Juan Fontana / Julie L Aspden / Abstract: Ribosomes have long been thought of as homogeneous macromolecular machines, but recent evidence suggests they are heterogeneous and could be specialised to regulate translation. Here, we have ...Ribosomes have long been thought of as homogeneous macromolecular machines, but recent evidence suggests they are heterogeneous and could be specialised to regulate translation. Here, we have characterised ribosomal protein heterogeneity across 4 tissues of Drosophila melanogaster. We find that testes and ovaries contain the most heterogeneous ribosome populations, which occurs through a combination of paralog-enrichment and paralog-switching. We have solved structures of ribosomes purified from in vivo tissues by cryo-EM, revealing differences in precise ribosomal arrangement for testis and ovary 80S ribosomes. Differences in the amino acid composition of paralog pairs and their localisation on the ribosome exterior indicate paralog-switching could alter the ribosome surface, enabling different proteins to regulate translation. One testis-specific paralog-switching pair is also found in humans, suggesting this is a conserved site of ribosome heterogeneity. Overall, this work allows us to propose that mRNA translation might be regulated in the gonads through ribosome heterogeneity, providing a potential means of ribosome specialisation. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 6xu6.cif.gz | 4.9 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb6xu6.ent.gz | Display | PDB format | |
| PDBx/mmJSON format | 6xu6.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/xu/6xu6ftp://data.pdbj.org/pub/pdb/validation_reports/xu/6xu6 | HTTPS FTP |
|---|
-Related structure data
| Related structure data | 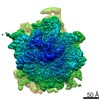 10622MC 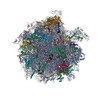 6xu7C 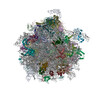 6xu8C M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |
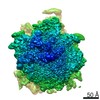

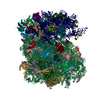
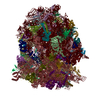
-Links
 PDBj
PDBj































- Assembly
Assembly
| Deposited unit | 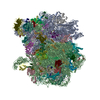
|
|---|---|
| 1 |
|
-Components
+40S ribosomal protein ... , 30 types, 30 molecules AAABACAUAXAMAdANALARAVAYAZAaAbADAeAJAEAGAHAIAQAKATAFAOAcAWAS
+60S ribosomal protein ... , 31 types, 31 molecules CACBCCCaCNCICDCQCRCSCPCZCrCbCeCiCkClCnCpCJCHCGCzCOCLCVCMCWCFCd
-Protein , 14 types, 14 molecules AgAPAfCTCXCYChCcCfCmCoDACgCj
| #7: Protein | Mass: 35661.934 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|---|
| #15: Protein | Mass: 14504.303 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #23: Protein | Mass: 9421.213 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #32: Protein | Mass: 18388.678 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #34: Protein | Mass: 13981.525 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #35: Protein | Mass: 15337.095 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #38: Protein | Mass: 14491.677 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #40: Protein | Mass: 11035.795 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #42: Protein | Mass: 17701.838 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #46: Protein | Mass: 6200.603 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #49: Protein | Mass: 12540.161 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #69: Protein | Mass: 41409.230 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #77: Protein | Mass: 12095.448 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #79: Protein | Mass: 9978.610 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
-Ribosomal protein ... , 2 types, 2 molecules CECU
| #52: Protein | Mass: 26179.066 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|---|
| #78: Protein | Mass: 11525.559 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
-RNA chain , 5 types, 5 molecules A9A7A8B2A5
| #54: RNA chain | Mass: 9665.705 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|---|
| #55: RNA chain | Mass: 38627.906 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #56: RNA chain | Mass: 39616.551 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #58: RNA chain | Mass: 641830.375 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #59: RNA chain | Mass: 1278930.125 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
-Details
| Has protein modification | Y |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: Drosophila melanogaster testis 80S ribosome / Type: RIBOSOME / Entity ID: all / Source: NATURAL |
|---|---|
| Molecular weight | Experimental value: NO |
| Source (natural) | Organism: |
| Buffer solution | pH: 7.5 |
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD |
| Image recording | Electron dose: 80 e/Å2 / Film or detector model: FEI FALCON II (4k x 4k) |
- Processing
Processing
| CTF correction | Type: PHASE FLIPPING ONLY |
|---|---|
| Symmetry | Point symmetry: C1 (asymmetric) |
| 3D reconstruction | Resolution: 3.5 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 46878 / Symmetry type: POINT |
| Atomic model building | Protocol: FLEXIBLE FIT |
| Atomic model building | PDB-ID: 4V6W Accession code: 4V6W / Source name: PDB / Type: experimental model |

