 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 6t64 | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | A model of the EIAV CA-SP hexamer (C6) from Gag-deltaMA spheres assembled at pH6 | |||||||||||||||||||||||||||||||||
 要素 要素 | Gag polyprotein | |||||||||||||||||||||||||||||||||
 キーワード キーワード | VIRAL PROTEIN / Retrovirus / lentivirus / Equine infectious anemia virus / EIAV / Gag / capsid / IP6 / phytic acid / inositolhexakiphosphate | |||||||||||||||||||||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報viral budding via host ESCRT complex / viral nucleocapsid / structural constituent of virion / nucleic acid binding / zinc ion binding 類似検索 - 分子機能 | |||||||||||||||||||||||||||||||||
| 生物種 |  Equine infectious anemia virus (ウマ伝染性貧血ウイルス) Equine infectious anemia virus (ウマ伝染性貧血ウイルス) | |||||||||||||||||||||||||||||||||
| 手法 | 電子顕微鏡法 / サブトモグラム平均法 / クライオ電子顕微鏡法 / 解像度: 3.7 Å | |||||||||||||||||||||||||||||||||
 データ登録者 データ登録者 | Dick, R.A. / Xu, C. / Morado, D.R. / Kravchuk, V. / Ricana, C.L. / Lyddon, T.D. / Broad, A.M. / Feathers, J.R. / Johnson, M.C. / Vogt, V.M. ...Dick, R.A. / Xu, C. / Morado, D.R. / Kravchuk, V. / Ricana, C.L. / Lyddon, T.D. / Broad, A.M. / Feathers, J.R. / Johnson, M.C. / Vogt, V.M. / Perilla, J.R. / Briggs, J.A.G. / Schur, F.K.M. | |||||||||||||||||||||||||||||||||
| 資金援助 |  オーストリア, オーストリア,  米国, 米国,  英国, 英国,  ドイツ, 10件 ドイツ, 10件
| |||||||||||||||||||||||||||||||||
 引用 引用 | ジャーナル: PLoS Pathog / 年: 2020 タイトル: Structures of immature EIAV Gag lattices reveal a conserved role for IP6 in lentivirus assembly. 著者: Robert A Dick / Chaoyi Xu / Dustin R Morado / Vladyslav Kravchuk / Clifton L Ricana / Terri D Lyddon / Arianna M Broad / J Ryan Feathers / Marc C Johnson / Volker M Vogt / Juan R Perilla / ...著者: Robert A Dick / Chaoyi Xu / Dustin R Morado / Vladyslav Kravchuk / Clifton L Ricana / Terri D Lyddon / Arianna M Broad / J Ryan Feathers / Marc C Johnson / Volker M Vogt / Juan R Perilla / John A G Briggs / Florian K M Schur / 要旨: Retrovirus assembly is driven by the multidomain structural protein Gag. Interactions between the capsid domains (CA) of Gag result in Gag multimerization, leading to an immature virus particle that ...Retrovirus assembly is driven by the multidomain structural protein Gag. Interactions between the capsid domains (CA) of Gag result in Gag multimerization, leading to an immature virus particle that is formed by a protein lattice based on dimeric, trimeric, and hexameric protein contacts. Among retroviruses the inter- and intra-hexamer contacts differ, especially in the N-terminal sub-domain of CA (CANTD). For HIV-1 the cellular molecule inositol hexakisphosphate (IP6) interacts with and stabilizes the immature hexamer, and is required for production of infectious virus particles. We have used in vitro assembly, cryo-electron tomography and subtomogram averaging, atomistic molecular dynamics simulations and mutational analyses to study the HIV-related lentivirus equine infectious anemia virus (EIAV). In particular, we sought to understand the structural conservation of the immature lentivirus lattice and the role of IP6 in EIAV assembly. Similar to HIV-1, IP6 strongly promoted in vitro assembly of EIAV Gag proteins into virus-like particles (VLPs), which took three morphologically highly distinct forms: narrow tubes, wide tubes, and spheres. Structural characterization of these VLPs to sub-4Å resolution unexpectedly showed that all three morphologies are based on an immature lattice with preserved key structural components, highlighting the structural versatility of CA to form immature assemblies. A direct comparison between EIAV and HIV revealed that both lentiviruses maintain similar immature interfaces, which are established by both conserved and non-conserved residues. In both EIAV and HIV-1, IP6 regulates immature assembly via conserved lysine residues within the CACTD and SP. Lastly, we demonstrate that IP6 stimulates in vitro assembly of immature particles of several other retroviruses in the lentivirus genus, suggesting a conserved role for IP6 in lentiviral assembly. | |||||||||||||||||||||||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 6t64.cif.gz | 135.7 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb6t64.ent.gz | 99.3 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 6t64.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/t6/6t64ftp://data.pdbj.org/pub/pdb/validation_reports/t6/6t64 | HTTPS FTP |
|---|
-関連構造データ








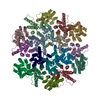

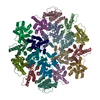


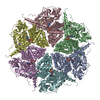
-リンク
 PDBj
PDBj
- 集合体
集合体
| 登録構造単位 | 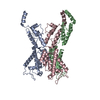
|
|---|---|
| 1 | x 6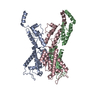
|
-要素
| #1: タンパク質 | 分子量: 54881.535 Da / 分子数: 3 / 由来タイプ: 組換発現 由来: (組換発現) Equine infectious anemia virus (ウマ伝染性貧血ウイルス)遺伝子: gag / 発現宿主:  Has protein modification | Y | |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: サブトモグラム平均法 |
- 試料調製
試料調製
| 構成要素 | 名称: Equine infectious anemia virus / タイプ: VIRUS 詳細: Gag construct was expressed in E.coli and purified using the SUMO-tag system. Assembly was performed at pH6. Entity ID: all / 由来: RECOMBINANT | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 実験値: NO | ||||||||||||||||||||
| 由来(天然) | 生物種: Equine infectious anemia virus (ウマ伝染性貧血ウイルス) | ||||||||||||||||||||
| 由来(組換発現) | 生物種: | ||||||||||||||||||||
| ウイルスについての詳細 | 中空か: YES / エンベロープを持つか: NO / 単離: OTHER / タイプ: VIRUS-LIKE PARTICLE | ||||||||||||||||||||
| 天然宿主 | 生物種: Equus caballus | ||||||||||||||||||||
| ウイルス殻 | 名称: Capsid / 直径: 1000 nm | ||||||||||||||||||||
| 緩衝液 | pH: 6 | ||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES 詳細: Virus-like-particles (spherical) of EIAV Gag deltaMAdeltap9 (referred to as Gag deltaMA) assembled at pH6. | ||||||||||||||||||||
| 試料支持 | 詳細: 20 mA / グリッドの材料: COPPER / グリッドのサイズ: 300 divisions/in. / グリッドのタイプ: C-flat-2/2 | ||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK II / 凍結剤: ETHANE / 湿度: 90 % / 凍結前の試料温度: 15 K / 詳細: 1-2 seconds blot time, offset -3mm |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS / 詳細: nanoprobe |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 130000 X / 最大 デフォーカス(公称値): -3500 nm / 最小 デフォーカス(公称値): -1500 nm / Cs: 2.7 mm / C2レンズ絞り径: 50 µm / アライメント法: ZEMLIN TABLEAU |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 平均露光時間: 1.4 sec. / 電子線照射量: 3.4 e/Å2 / 検出モード: SUPER-RESOLUTION フィルム・検出器のモデル: GATAN K2 QUANTUM (4k x 4k) 撮影したグリッド数: 1 詳細: Data was acquired using a dose-symmetric tilt acquisition scheme, as described in Hagen et al, 2017, J. Struct. Biol, 197(2):191-8 |
| 電子光学装置 | エネルギーフィルター名称: GIF Quantum LS / エネルギーフィルタースリット幅: 20 eV |
| 画像スキャン | 横: 3708 / 縦: 3838 / 動画フレーム数/画像: 21 |
- 解析
解析
| ソフトウェア | 名称: PHENIX / バージョン: 1.14_3260: / 分類: 精密化 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 画像処理 | 詳細: Tilt series were low-pass filtered according to their cumulative dose using exposure filters that were calculated using an exposure-dependent amplitude attenuation function and critical ...詳細: Tilt series were low-pass filtered according to their cumulative dose using exposure filters that were calculated using an exposure-dependent amplitude attenuation function and critical exposure constants (as published in Grant & Grigorieff, Elife, 2015). Tilt series were aligned and reconstructed in IMOD. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CTF補正 | 詳細: CTF-correction was performed using NOVACTF / タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C6 (6回回転対称) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.7 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 64961 / クラス平均像の数: 1 / 対称性のタイプ: POINT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EM volume selection | 手法: Subvolumes were defined according to their position on the VLPs 詳細: Subtomogram extraction positions were defined in Amira using the electron microscopy toolbox by determing the radii and the center of the VLPs. Initially, positions were oversampled and ...詳細: Subtomogram extraction positions were defined in Amira using the electron microscopy toolbox by determing the radii and the center of the VLPs. Initially, positions were oversampled and subsequently cleaned during alignments using cross-correlation and distance thresholds. Num. of tomograms: 55 / Num. of volumes extracted: 311207 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: OTHER / 空間: REAL / Target criteria: Cross-correlation coefficient 詳細: Rigid body fitting was done in Chimera. Missing residues were built de novo in Coot. Refinement was performed iteratively in Phenix and Coot. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | PDB-ID: 2EIA Accession code: 2EIA / Source name: PDB / タイプ: experimental model |

