 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 6c24 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
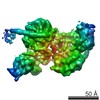
| タイトル | Cryo-EM structure of PRC2 bound to cofactors AEBP2 and JARID2 in the Extended Active State | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | GENE REGULATION / polycomb repressive complex / AEBP2 / JARID2 / histone modification | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報protein localization to pericentric heterochromatin / regulation of kidney development / hepatocyte homeostasis / cellular response to trichostatin A / regulation of gliogenesis / [histone H3]-lysine27 N-trimethyltransferase / negative regulation of striated muscle cell differentiation / regulatory ncRNA-mediated heterochromatin formation / sex chromatin / CAF-1 complex ...protein localization to pericentric heterochromatin / regulation of kidney development / hepatocyte homeostasis / cellular response to trichostatin A / regulation of gliogenesis / [histone H3]-lysine27 N-trimethyltransferase / negative regulation of striated muscle cell differentiation / regulatory ncRNA-mediated heterochromatin formation / sex chromatin / CAF-1 complex / negative regulation of keratinocyte differentiation / histone H3K27 trimethyltransferase activity / negative regulation of retinoic acid receptor signaling pathway / random inactivation of X chromosome / primary miRNA binding / ubiquitin-modified histone reader activity / skeletal muscle satellite cell maintenance involved in skeletal muscle regeneration / response to tetrachloromethane / cerebellar cortex development / facultative heterochromatin formation / histone H3K27 methyltransferase activity / negative regulation of cardiac muscle hypertrophy / negative regulation of cardiac muscle cell proliferation / positive regulation of cell cycle G1/S phase transition / NURF complex / regulation of cell fate specification / negative regulation of stem cell population maintenance / DNA replication-dependent chromatin assembly / chromatin silencing complex / Transcription of E2F targets under negative control by p107 (RBL1) and p130 (RBL2) in complex with HDAC1 / regulation of stem cell differentiation / NuRD complex / ESC/E(Z) complex / protein-lysine N-methyltransferase activity / negative regulation of stem cell differentiation / Transcription of E2F targets under negative control by DREAM complex / pronucleus / RSC-type complex / cardiac muscle hypertrophy in response to stress / Polo-like kinase mediated events / synaptic transmission, GABAergic / lncRNA binding / positive regulation of dendrite development / histone H3 methyltransferase activity / cardiac muscle cell proliferation / histone methyltransferase complex / negative regulation of G1/S transition of mitotic cell cycle / negative regulation of gene expression, epigenetic / spinal cord development / G1 to G0 transition / Sin3-type complex / histone methyltransferase activity / positive regulation of stem cell population maintenance / G1/S-Specific Transcription / ATPase complex / oligodendrocyte differentiation / negative regulation of transcription elongation by RNA polymerase II / Transcriptional Regulation by E2F6 / RNA Polymerase I Transcription Initiation / negative regulation of cell differentiation / histone deacetylase complex / G0 and Early G1 / subtelomeric heterochromatin formation / negative regulation of cytokine production involved in inflammatory response / RNA polymerase II core promoter sequence-specific DNA binding / pericentric heterochromatin / ribonucleoprotein complex binding / positive regulation of epithelial to mesenchymal transition / Cyclin E associated events during G1/S transition / Cyclin A:Cdk2-associated events at S phase entry / nucleosome binding / keratinocyte differentiation / Deposition of new CENPA-containing nucleosomes at the centromere / heterochromatin formation / spleen development / protein localization to chromatin / enzyme activator activity / Regulation of TP53 Activity through Acetylation / methylated histone binding / SUMOylation of chromatin organization proteins / negative regulation of cell migration / B cell differentiation / transcription corepressor binding / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / positive regulation of GTPase activity / thymus development / liver development / PRC2 methylates histones and DNA / ubiquitin binding / Regulation of PTEN gene transcription / cellular response to leukemia inhibitory factor / Defective pyroptosis / central nervous system development / HDACs deacetylate histones / stem cell differentiation / hippocampus development / liver regeneration / promoter-specific chromatin binding / transcription coregulator activity / negative regulation of transforming growth factor beta receptor signaling pathway 類似検索 - 分子機能 | ||||||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.5 Å | ||||||||||||
 データ登録者 データ登録者 | Kasinath, V. / Faini, M. / Poepsel, S. / Reif, D. / Feng, A. / Stjepanovic, G. / Aebersold, R. / Nogales, E. | ||||||||||||
| 資金援助 |  米国, European Union, 3件 米国, European Union, 3件
| ||||||||||||
 引用 引用 | ジャーナル: Science / 年: 2018 タイトル: Structures of human PRC2 with its cofactors AEBP2 and JARID2. 著者: Vignesh Kasinath / Marco Faini / Simon Poepsel / Dvir Reif / Xinyu Ashlee Feng / Goran Stjepanovic / Ruedi Aebersold / Eva Nogales /  要旨: Transcriptionally repressive histone H3 lysine 27 methylation by Polycomb repressive complex 2 (PRC2) is essential for cellular differentiation and development. Here we report cryo-electron ...Transcriptionally repressive histone H3 lysine 27 methylation by Polycomb repressive complex 2 (PRC2) is essential for cellular differentiation and development. Here we report cryo-electron microscopy structures of human PRC2 in a basal state and two distinct active states while in complex with its cofactors JARID2 and AEBP2. Both cofactors mimic the binding of histone H3 tails. JARID2, methylated by PRC2, mimics a methylated H3 tail to stimulate PRC2 activity, whereas AEBP2 interacts with the RBAP48 subunit, mimicking an unmodified H3 tail. SUZ12 interacts with all other subunits within the assembly and thus contributes to the stability of the complex. Our analysis defines the complete architecture of a functionally relevant PRC2 and provides a structural framework to understand its regulation by cofactors, histone tails, and RNA. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 6c24.cif.gz | 418.6 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb6c24.ent.gz | 301.8 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 6c24.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 6c24_validation.pdf.gz | 1.2 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 6c24_full_validation.pdf.gz | 1.3 MB | 表示 | |
| XML形式データ | 6c24_validation.xml.gz | 63.9 KB | 表示 | |
| CIF形式データ | 6c24_validation.cif.gz | 96.1 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/c2/6c24ftp://data.pdbj.org/pub/pdb/validation_reports/c2/6c24 | HTTPS FTP |
-関連構造データ




-リンク
 PDBj
PDBj



















- 集合体
集合体
| 登録構造単位 | 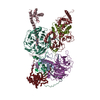
|
|---|---|
| 1 |
|
-要素
-Polycomb protein ... , 2種, 4分子 AMQL
| #1: タンパク質 | 分子量: 83181.922 Da / 分子数: 3 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: SUZ12, CHET9, JJAZ1, KIAA0160 / 発現宿主:  Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q15022 Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q15022#5: タンパク質 | | 分子量: 50267.691 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: EED / 発現宿主: Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: O75530 |
|---|
-タンパク質 , 6種, 7分子 BEKCNPZ
| #2: タンパク質 | 分子量: 37706.449 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: JARID2, JMJ / 発現宿主: Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q92833 | ||||||
|---|---|---|---|---|---|---|---|
| #3: タンパク質 | 分子量: 37664.371 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: JARID2, JMJ / 発現宿主: Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q92833 | ||||||
| #4: タンパク質 | 分子量: 85492.297 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: EZH2, KMT6 / 発現宿主: Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q15910, histone-lysine N-methyltransferase#6: タンパク質 | | 分子量: 47709.527 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: RBBP4, RBAP48 / 発現宿主: Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q09028#8: タンパク質 | | 分子量: 33012.668 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: AEBP2 / 発現宿主: Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q6ZN18#9: タンパク質 | | 分子量: 11507.176 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 発現宿主: Trichoplusia ni (イラクサキンウワバ) |
-タンパク質・ペプチド , 1種, 1分子 O
| #7: タンパク質・ペプチド | 分子量: 735.874 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 発現宿主: Trichoplusia ni (イラクサキンウワバ) |
|---|
-詳細
| 配列の詳細 | authors have indicated that the macromolecule name is corresponding to chain Z is SUZ12. The ...authors have indicated that the macromolecule name is corresponding to chain Z is SUZ12. The expected sequence for this portion is: SUZ12 aa 150 - 356 SHSLSAHLQL |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Ternary complex of PRC2 with cofactors AEBP2 and JARID2 タイプ: COMPLEX / Entity ID: all / 由来: RECOMBINANT |
|---|---|
| 分子量 | 値: 0.3 MDa / 実験値: NO |
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 由来(組換発現) | 生物種: Trichoplusia ni (イラクサキンウワバ) |
| 緩衝液 | pH: 7.9 |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 急速凍結 | 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: SPOT SCAN FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: SPOT SCAN |
| 電子レンズ | モード: BRIGHT FIELD |
| 撮影 | 電子線照射量: 60 e/Å2 / 検出モード: SUPER-RESOLUTION フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) |
| 電子光学装置 | 位相板: Volta Phase Plate |
- 解析
解析
| ソフトウェア | 名称: REFMAC / バージョン: 5.8.0088 / 分類: 精密化 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING ONLY | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.5 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 249696 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 | 解像度: 3.5→151.2 Å / Cor.coef. Fo:Fc: 0.843 / SU B: 29.608 / SU ML: 0.428 / ESU R: 0.808 立体化学のターゲット値: MAXIMUM LIKELIHOOD WITH PHASES 詳細: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | イオンプローブ半径: 0.8 Å / 減衰半径: 0.8 Å / VDWプローブ半径: 1.2 Å / 溶媒モデル: MASK | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 114.285 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: 1 / 合計: 13350 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
|