ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
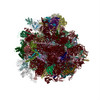
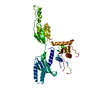
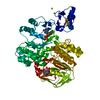
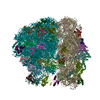
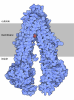
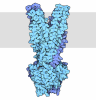
| 登録情報 | データベース: PDB / ID: 3jai | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | Structure of a mammalian ribosomal termination complex with ABCE1, eRF1(AAQ), and the UGA stop codon | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | RIBOSOME / termination / eRF1 / ABCE1 | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報translation termination factor activity / translation release factor complex / cytoplasmic translational termination / translation release factor activity / regulation of translational termination / ribosomal subunit / protein methylation / translation release factor activity, codon specific / sequence-specific mRNA binding / nuclear-transcribed mRNA catabolic process, nonsense-mediated decay ...translation termination factor activity / translation release factor complex / cytoplasmic translational termination / translation release factor activity / regulation of translational termination / ribosomal subunit / protein methylation / translation release factor activity, codon specific / sequence-specific mRNA binding / nuclear-transcribed mRNA catabolic process, nonsense-mediated decay / aminoacyl-tRNA hydrolase activity / regulation of G1 to G0 transition / positive regulation of intrinsic apoptotic signaling pathway in response to DNA damage by p53 class mediator / regulation of translation involved in cellular response to UV / protein-DNA complex disassembly / positive regulation of DNA damage response, signal transduction by p53 class mediator resulting in transcription of p21 class mediator / mammalian oogenesis stage / G1 to G0 transition / activation-induced cell death of T cells / Protein hydroxylation / positive regulation of signal transduction by p53 class mediator / ubiquitin ligase inhibitor activity / phagocytic cup / Eukaryotic Translation Termination / Nonsense Mediated Decay (NMD) independent of the Exon Junction Complex (EJC) / ribosomal small subunit binding / endonucleolytic cleavage to generate mature 3'-end of SSU-rRNA from (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / TOR signaling / T cell proliferation involved in immune response / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / ribosomal small subunit export from nucleus / erythrocyte development / translation regulator activity / cellular response to actinomycin D / ribosomal subunit export from nucleus / cytosolic ribosome / translational termination / rough endoplasmic reticulum / endonucleolytic cleavage in ITS1 to separate SSU-rRNA from 5.8S rRNA and LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / gastrulation / MDM2/MDM4 family protein binding / translational initiation / DNA damage response, signal transduction by p53 class mediator resulting in cell cycle arrest / class I DNA-(apurinic or apyrimidinic site) endonuclease activity / DNA-(apurinic or apyrimidinic site) lyase / negative regulation of ubiquitin-dependent protein catabolic process / rescue of stalled ribosome / maturation of LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / 90S preribosome / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / maturation of LSU-rRNA / ribosomal large subunit biogenesis / cellular response to leukemia inhibitory factor / maturation of SSU-rRNA / positive regulation of translation / small-subunit processome / positive regulation of apoptotic signaling pathway / protein kinase C binding / positive regulation of protein-containing complex assembly / placenta development / cellular response to gamma radiation / mRNA 5'-UTR binding / Regulation of expression of SLITs and ROBOs / transcription coactivator binding / cytoplasmic ribonucleoprotein granule / modification-dependent protein catabolic process / spindle / G1/S transition of mitotic cell cycle / rRNA processing / protein tag activity / rhythmic process / positive regulation of canonical Wnt signaling pathway / ribosome biogenesis / ribosome binding / glucose homeostasis / regulation of translation / ribosomal small subunit biogenesis / ribosomal small subunit assembly / small ribosomal subunit / small ribosomal subunit rRNA binding / cell body / T cell differentiation in thymus / 5S rRNA binding / large ribosomal subunit rRNA binding / perikaryon / cytosolic small ribosomal subunit / cytosolic large ribosomal subunit / mitochondrial inner membrane / tRNA binding / cytoplasmic translation / postsynaptic density / cell differentiation / protein stabilization / rRNA binding / ribosome / protein ubiquitination / structural constituent of ribosome / ribonucleoprotein complex / iron ion binding / translation 類似検索 - 分子機能 | ||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.65 Å | ||||||
 データ登録者 データ登録者 | Brown, A. / Shao, S. / Murray, J. / Hegde, R.S. / Ramakrishnan, V. | ||||||
 引用 引用 | ジャーナル: Nature / 年: 2015 タイトル: Structural basis for stop codon recognition in eukaryotes. 著者: Alan Brown / Sichen Shao / Jason Murray / Ramanujan S Hegde / V Ramakrishnan /  要旨: Termination of protein synthesis occurs when a translating ribosome encounters one of three universally conserved stop codons: UAA, UAG or UGA. Release factors recognize stop codons in the ribosomal ...Termination of protein synthesis occurs when a translating ribosome encounters one of three universally conserved stop codons: UAA, UAG or UGA. Release factors recognize stop codons in the ribosomal A-site to mediate release of the nascent chain and recycling of the ribosome. Bacteria decode stop codons using two separate release factors with differing specificities for the second and third bases. By contrast, eukaryotes rely on an evolutionarily unrelated omnipotent release factor (eRF1) to recognize all three stop codons. The molecular basis of eRF1 discrimination for stop codons over sense codons is not known. Here we present cryo-electron microscopy (cryo-EM) structures at 3.5-3.8 Å resolution of mammalian ribosomal complexes containing eRF1 interacting with each of the three stop codons in the A-site. Binding of eRF1 flips nucleotide A1825 of 18S ribosomal RNA so that it stacks on the second and third stop codon bases. This configuration pulls the fourth position base into the A-site, where it is stabilized by stacking against G626 of 18S rRNA. Thus, eRF1 exploits two rRNA nucleotides also used during transfer RNA selection to drive messenger RNA compaction. In this compacted mRNA conformation, stop codons are favoured by a hydrogen-bonding network formed between rRNA and essential eRF1 residues that constrains the identity of the bases. These results provide a molecular framework for eukaryotic stop codon recognition and have implications for future studies on the mechanisms of canonical and premature translation termination. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 3jai.cif.gz | 5.5 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb3jai.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 3jai.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 3jai_validation.pdf.gz | 1.5 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 3jai_full_validation.pdf.gz | 1.8 MB | 表示 | |
| XML形式データ | 3jai_validation.xml.gz | 369.1 KB | 表示 | |
| CIF形式データ | 3jai_validation.cif.gz | 644.5 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/ja/3jaiftp://data.pdbj.org/pub/pdb/validation_reports/ja/3jai | HTTPS FTP |
-関連構造データ




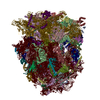
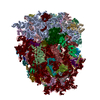

-リンク
 PDBj
PDBj




































- 集合体
集合体
| 登録構造単位 | 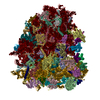
|
|---|---|
| 1 |
|
-要素
+タンパク質 , 77種, 77分子 ABCDEFGHIJLMNOPQRSTUVWXYZabcde...

-タンパク質・ペプチド , 3種, 3分子 ln1
| #37: タンパク質・ペプチド | 分子量: 6295.562 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #39: タンパク質・ペプチド | 分子量: 3213.075 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #45: タンパク質・ペプチド | 分子量: 1788.032 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-RNA鎖 , 7種, 7分子 235789hh
| #46: RNA鎖 | 分子量: 24436.551 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #47: RNA鎖 | 分子量: 24102.275 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #48: RNA鎖 | 分子量: 1186579.500 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #49: RNA鎖 | 分子量: 38691.914 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #50: RNA鎖 | 分子量: 50143.648 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #51: RNA鎖 | 分子量: 554751.312 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #85: RNA鎖 | 分子量: 3837.328 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-非ポリマー , 4種, 207分子 

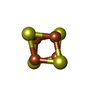

| #88: 化合物 | ChemComp-MG /  分子量: 24.305 Da / 分子数: 195 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 195 / 由来タイプ: 合成 / 式: Mg#89: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn#90: 化合物 |  分子量: 351.640 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Fe4S4 分子量: 351.640 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Fe4S4#91: 化合物 |  分子量: 427.201 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / コメント: ADP, エネルギー貯蔵分子*YM 分子量: 427.201 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / コメント: ADP, エネルギー貯蔵分子*YM |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 緩衝液 | 名称: 50 mM HEPES, 100 mM potassium acetate, 5 mM magnesium acetate, 1 mM DTT pH: 7.4 詳細: 50 mM HEPES, 100 mM potassium acetate, 5 mM magnesium acetate, 1 mM DTT | ||||||||||||||||||||||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||||||||||||
| 試料支持 | 詳細: Quantifoil R2/2 400 mesh Cu grid with thin continuous carbon support, glow discharged | ||||||||||||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK III / 凍結剤: ETHANE / 湿度: 100 % 詳細: After 30 second wait time, blot for 3 seconds before plunging into liquid ethane (FEI VITROBOT MARK III). 手法: After 30 second wait time, blot for 3 seconds before plunging |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS / 日付: 2015年4月17日 / 詳細: Automated data acquisition using EPU (FEI) |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 59000 X / 倍率(補正後): 104478 X / 最大 デフォーカス(公称値): 3600 nm / 最小 デフォーカス(公称値): 1700 nm / Cs: 2.7 mm |
| 試料ホルダ | 資料ホルダタイプ: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 電子線照射量: 30 e/Å2 フィルム・検出器のモデル: FEI FALCON II (4k x 4k) |
| 電子光学装置 | エネルギーフィルター名称: FEI |
| 画像スキャン | デジタル画像の数: 1472 |
| 放射波長 | 相対比: 1 |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||
| 3次元再構成 | 解像度: 3.65 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 22058 / ピクセルサイズ(公称値): 1.34 Å / ピクセルサイズ(実測値): 1.34 Å / 対称性のタイプ: POINT | ||||||||||||||||||||
| 原子モデル構築 |
| ||||||||||||||||||||
| 原子モデル構築 |
| ||||||||||||||||||||
| 精密化ステップ | サイクル: LAST
|