 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 3j46 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Structure of the SecY protein translocation channel in action | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード | RIBOSOME/PROTEIN TRANSPORT / 70S / preprotein translocase / SECYEG / protein translocation channel / nascent chain / RIBOSOME-PROTEIN TRANSPORT complex | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報protein insertion into membrane from inner side / cell envelope Sec protein transport complex / intracellular protein transmembrane transport / protein transport by the Sec complex / protein-transporting ATPase activity / SRP-dependent cotranslational protein targeting to membrane, translocation / signal sequence binding / protein secretion / protein transmembrane transporter activity / negative regulation of translational initiation ...protein insertion into membrane from inner side / cell envelope Sec protein transport complex / intracellular protein transmembrane transport / protein transport by the Sec complex / protein-transporting ATPase activity / SRP-dependent cotranslational protein targeting to membrane, translocation / signal sequence binding / protein secretion / protein transmembrane transporter activity / negative regulation of translational initiation / intracellular protein transport / ribosomal large subunit assembly / large ribosomal subunit rRNA binding / cytosolic large ribosomal subunit / cytoplasmic translation / tRNA binding / rRNA binding / structural constituent of ribosome / translation / membrane / plasma membrane / cytosol / cytoplasm 類似検索 - 分子機能 | |||||||||
| 生物種 |  | |||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 10.1 Å | |||||||||
 データ登録者 データ登録者 | Akey, C.W. / Park, E. / Menetret, J.F. / Gumbart, J.C. / Ludtke, S.J. / Li, W. / Whynot, A. / Rapoport, T.A. | |||||||||
 引用 引用 | ジャーナル: Nature / 年: 2014 タイトル: Structure of the SecY channel during initiation of protein translocation. 著者: Eunyong Park / Jean-François Ménétret / James C Gumbart / Steven J Ludtke / Weikai Li / Andrew Whynot / Tom A Rapoport / Christopher W Akey /  要旨: Many secretory proteins are targeted by signal sequences to a protein-conducting channel, formed by prokaryotic SecY or eukaryotic Sec61 complexes, and are translocated across the membrane during ...Many secretory proteins are targeted by signal sequences to a protein-conducting channel, formed by prokaryotic SecY or eukaryotic Sec61 complexes, and are translocated across the membrane during their synthesis. Crystal structures of the inactive channel show that the SecY subunit of the heterotrimeric complex consists of two halves that form an hourglass-shaped pore with a constriction in the middle of the membrane and a lateral gate that faces the lipid phase. The closed channel has an empty cytoplasmic funnel and an extracellular funnel that is filled with a small helical domain, called the plug. During initiation of translocation, a ribosome-nascent chain complex binds to the SecY (or Sec61) complex, resulting in insertion of the nascent chain. However, the mechanism of channel opening during translocation is unclear. Here we have addressed this question by determining structures of inactive and active ribosome-channel complexes with cryo-electron microscopy. Non-translating ribosome-SecY channel complexes derived from Methanocaldococcus jannaschii or Escherichia coli show the channel in its closed state, and indicate that ribosome binding per se causes only minor changes. The structure of an active E. coli ribosome-channel complex demonstrates that the nascent chain opens the channel, causing mostly rigid body movements of the amino- and carboxy-terminal halves of SecY. In this early translocation intermediate, the polypeptide inserts as a loop into the SecY channel with the hydrophobic signal sequence intercalated into the open lateral gate. The nascent chain also forms a loop on the cytoplasmic surface of SecY rather than entering the channel directly. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 3j46.cif.gz | 439.1 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb3j46.ent.gz | 305.7 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 3j46.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 3j46_validation.pdf.gz | 1.1 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 3j46_full_validation.pdf.gz | 1.4 MB | 表示 | |
| XML形式データ | 3j46_validation.xml.gz | 70.4 KB | 表示 | |
| CIF形式データ | 3j46_validation.cif.gz | 104.1 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/j4/3j46ftp://data.pdbj.org/pub/pdb/validation_reports/j4/3j46 | HTTPS FTP |
-関連構造データ




-リンク
 PDBj
PDBj





























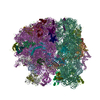
- 集合体
集合体
| 登録構造単位 | 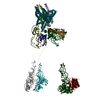
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 4種, 4分子 yEGn
| #1: タンパク質 | 分子量: 47659.371 Da / 分子数: 1 / 変異: S68C / 由来タイプ: 組換発現 / 由来: (組換発現) |
|---|---|
| #2: タンパク質 | 分子量: 6123.306 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) |
| #3: タンパク質 | 分子量: 6566.800 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) |
| #4: タンパク質 | 分子量: 10780.095 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) |
-RNA鎖 , 2種, 2分子 pa
| #5: RNA鎖 | 分子量: 24478.502 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #6: RNA鎖 | 分子量: 24599.748 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-50S ribosomal protein ... , 4種, 4分子 5TUY
| #7: タンパク質 | 分子量: 24765.660 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #8: タンパク質 | 分子量: 11222.160 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #9: タンパク質 | 分子量: 11208.054 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #10: タンパク質 | 分子量: 7286.464 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-23S ribosomal ... , 4種, 4分子 1234
| #11: RNA鎖 | 分子量: 20350.104 Da / 分子数: 1 / Fragment: helix 6 - helix 7 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #12: RNA鎖 | 分子量: 11661.961 Da / 分子数: 1 / Fragment: helix 50 / 由来タイプ: 天然 / 由来: (天然) |
| #13: RNA鎖 | 分子量: 14261.592 Da / 分子数: 1 / Fragment: helix 59 / 由来タイプ: 天然 / 由来: (天然) |
| #14: RNA鎖 | 分子量: 35116.715 Da / 分子数: 1 / Fragment: helix 76 - helix 78 / 由来タイプ: 天然 / 由来: (天然) |
-詳細
| Has protein modification | Y |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 / 使用した結晶の数: 1 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 値: 2.5 MDa / 実験値: NO | ||||||||||||||||||||
| 緩衝液 | 名称: 50 mM Tris-acetate, 10 mM Mg(OAc)2, 80 mM KOAc, 0.06% DDM pH: 7.2 詳細: 50 mM Tris-acetate, 10 mM Mg(OAc)2, 80 mM KOAc, 0.06% DDM | ||||||||||||||||||||
| 試料 | 濃度: 8 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||
| 試料支持 | 詳細: 400 mesh Quantifoil holey grids with 2/1 or 1.2/1.2 | ||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK III / 凍結剤: ETHANE / Temp: 77 K / 湿度: 95 % 詳細: Blot 1-2 seconds before plunging into liquid ethane (FEI VITROBOT MARK III). |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Tecnai F20 / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TECNAI F20 / 日付: 2012年2月10日 詳細: Low dose imaging: automated single particle data collection program from TVIPS was used. |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 160 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 160 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 42000 X / 最大 デフォーカス(公称値): 3000 nm / 最小 デフォーカス(公称値): 1000 nm / Cs: 2 mm |
| 試料ホルダ | 試料ホルダーモデル: OTHER / 資料ホルダタイプ: CT3500 / 温度: 94 K |
| 撮影 | 電子線照射量: 20 e/Å2 / フィルム・検出器のモデル: KODAK SO-163 FILM / 詳細: Tietz 4096 x 4096 CCD |
| 画像スキャン | デジタル画像の数: 4900 |
| 放射波長 | 相対比: 1 |
- 解析
解析
| ソフトウェア |
| ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||||||||
| CTF補正 | 詳細: per micrograph | ||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||||||||
| 3次元再構成 | 手法: projection matching / 解像度: 10.1 Å / 解像度の算出法: FSC 0.5 CUT-OFF / 粒子像の数: 53000 / ピクセルサイズ(公称値): 2.12 Å / ピクセルサイズ(実測値): 2.12 Å 詳細: The structure was solved twice: first with a model starting from a 25-Angstrom filtered E. coli ribosome map generated in house, and then a second time using a filtered ribosome model (EMD- ...詳細: The structure was solved twice: first with a model starting from a 25-Angstrom filtered E. coli ribosome map generated in house, and then a second time using a filtered ribosome model (EMD-5036). In each case, after convergence, maps from two EMAN2 refinements with different parameters were averaged after alignment in Chimera. Four maps in total were averaged to reduce the noise. Resolution method was FSC at 0.5 cut-off for a comparison between the full experimental 3D density map and a calculated map of the docked E. coli ribosome model (this map was calculated to 7 Angstrom resolution with EMAN). クラス平均像の数: 5300 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||
| 原子モデル構築 |
| ||||||||||||||||||||||||||||||
| 原子モデル構築 | Source name: PDB / タイプ: experimental model
| ||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: LAST
|

