 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: EMDB / ID: EMD-9032 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
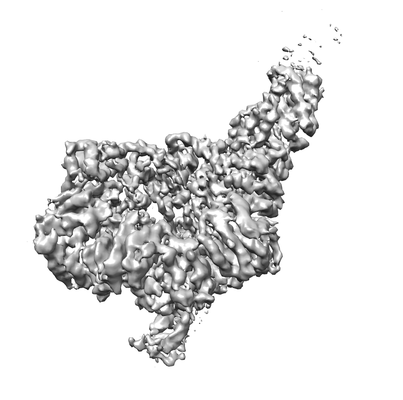
| タイトル | Full-length S. pombe Mdn1 in the presence of AMPPNP (ring region) | |||||||||
 マップデータ マップデータ | Full-length S. pombe Mdn1 in the presence of AMPPNP (ring region), primary map | |||||||||
 試料 試料 |
| |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報preribosome, large subunit precursor / ribosomal large subunit export from nucleus / ribosomal large subunit biogenesis / ribosome biogenesis / ribosomal large subunit assembly / calcium ion binding / nucleolus / ATP hydrolysis activity / nucleoplasm / ATP binding / nucleus 類似検索 - 分子機能 | |||||||||
| 生物種 |  | |||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 4.0 Å | |||||||||
 データ登録者 データ登録者 | Chen Z / Suzuki H / Wang AC / DiMaio F / Walz T / Kapoor TM | |||||||||
 引用 引用 | ジャーナル: Cell / 年: 2018 タイトル: Structural Insights into Mdn1, an Essential AAA Protein Required for Ribosome Biogenesis. 著者: Zhen Chen / Hiroshi Suzuki / Yuki Kobayashi / Ashley C Wang / Frank DiMaio / Shigehiro A Kawashima / Thomas Walz / Tarun M Kapoor /   要旨: Mdn1 is an essential AAA (ATPase associated with various activities) protein that removes assembly factors from distinct precursors of the ribosomal 60S subunit. However, Mdn1's large size (∼5,000 ...Mdn1 is an essential AAA (ATPase associated with various activities) protein that removes assembly factors from distinct precursors of the ribosomal 60S subunit. However, Mdn1's large size (∼5,000 amino acid [aa]) and its limited homology to other well-studied proteins have restricted our understanding of its remodeling function. Here, we present structures for S. pombe Mdn1 in the presence of AMPPNP at up to ∼4 Å or ATP plus Rbin-1, a chemical inhibitor, at ∼8 Å resolution. These data reveal that Mdn1's MIDAS domain is tethered to its ring-shaped AAA domain through an ∼20 nm long structured linker and a flexible ∼500 aa Asp/Glu-rich motif. We find that the MIDAS domain, which also binds other ribosome-assembly factors, docks onto the AAA ring in a nucleotide state-specific manner. Together, our findings reveal how conformational changes in the AAA ring can be directly transmitted to the MIDAS domain and thereby drive the targeted release of assembly factors from ribosomal 60S-subunit precursors. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | EMマップ: SurfViewMolmilJmol/JSmol |
| 添付画像 |
- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_9032.map.gz | 91.8 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-9032-v30.xmlemd-9032.xml | 17 KB 17 KB | 表示 表示 | EMDBヘッダ |
| FSC (解像度算出) | emd_9032_fsc.xml | 8.7 KB | 表示 | FSCデータファイル |
| 画像 |  emd_9032.png emd_9032.png | 45.5 KB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-9032ftp://ftp.pdbj.org/pub/emdb/structures/EMD-9032 http://ftp.pdbj.org/pub/emdb/structures/EMD-9032ftp://ftp.pdbj.org/pub/emdb/structures/EMD-9032 | HTTPS FTP |
-検証レポート
| 文書・要旨 | emd_9032_validation.pdf.gz | 489.7 KB | 表示 | EMDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | emd_9032_full_validation.pdf.gz | 489.3 KB | 表示 | |
| XML形式データ | emd_9032_validation.xml.gz | 11.3 KB | 表示 | |
| CIF形式データ | emd_9032_validation.cif.gz | 14.8 KB | 表示 | |
| アーカイブディレクトリ | https://ftp.pdbj.org/pub/emdb/validation_reports/EMD-9032ftp://ftp.pdbj.org/pub/emdb/validation_reports/EMD-9032 | HTTPS FTP |
-関連構造データ





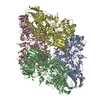
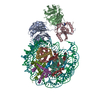
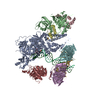


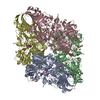
-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| 「今月の分子」の関連する項目 |



-マップ
| ファイル | ダウンロード / ファイル: emd_9032.map.gz / 形式: CCP4 / 大きさ: 103 MB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | Full-length S. pombe Mdn1 in the presence of AMPPNP (ring region), primary map | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 投影像・断面図 | 画像のコントロール
画像は Spider により作成 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ボクセルのサイズ | X=Y=Z: 1.3 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 密度 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 詳細 | EMDB XML:
CCP4マップ ヘッダ情報:
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-添付データ
- 試料の構成要素
試料の構成要素
-全体 : N-terminal density map of the full-length Mdn1 in the presence of...
| 全体 | 名称: N-terminal density map of the full-length Mdn1 in the presence of AMPPNP |
|---|---|
| 要素 |
|
-超分子 #1: N-terminal density map of the full-length Mdn1 in the presence of...
| 超分子 | 名称: N-terminal density map of the full-length Mdn1 in the presence of AMPPNP タイプ: organelle_or_cellular_component / ID: 1 / 親要素: 0 / 含まれる分子: #1 |
|---|---|
| 由来(天然) | 生物種: |
| 分子量 | 理論値: 540 KDa |
| 組換発現 | 生物種:  Trichoplusia ni (イラクサキンウワバ) Trichoplusia ni (イラクサキンウワバ) |
-分子 #1: Midasin
| 分子 | 名称: Midasin / タイプ: protein_or_peptide / ID: 1 / コピー数: 1 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: 株: 972 / ATCC 24843 |
| 分子量 | 理論値: 262.860312 KDa |
| 組換発現 | 生物種: Trichoplusia ni (イラクサキンウワバ) |
| 配列 | 文字列: MDVLIEWVAI YPQIYDILEH INYVPSNTLQ RLRLHQPWSK IDYDVWFLYA SDEIRETCKV KYYGETKTYG EVFVLENERI SQLHRLFVS WTVSERAEHL KNLLFDAGLS NLPLVELGGN VFFNSHVPLP CSLVLTKSTQ ENLNRITPYL VQKRPILLAG P EGIGKKFL ...文字列: MDVLIEWVAI YPQIYDILEH INYVPSNTLQ RLRLHQPWSK IDYDVWFLYA SDEIRETCKV KYYGETKTYG EVFVLENERI SQLHRLFVS WTVSERAEHL KNLLFDAGLS NLPLVELGGN VFFNSHVPLP CSLVLTKSTQ ENLNRITPYL VQKRPILLAG P EGIGKKFL ITQIAAKLGQ QIIRIHLSDS TDPKMLIGTY TSPKPGEFEW QPGVLTQAVI TGKWILFTNI EHAPSEVLSV LL PLLEKRQ LVIPSRGETI YAKGSFQMFA TSSMKTKILG QRLWQILDLT YQPDECVEVV STLYPVLSII CPTLYSVYKD IFD LFSQRS FLATSKIYRR LCLRDFYKFI KRVAFLYHKF MIPSDHVVIS QELQDAVFKE AIDMFGAFIP SRDGFDLVVR NVAI ELNIP PEKALQLRYS IPVFQNLEHN INIGRCSLKK LSTIRSCSTN SYAFTSSSLG LLEQLAAGVQ TNEPLLLVGE TGTGK TTTI QLLAGLLGQK VTVINMSQQT ESSDMLGGYK PINASTLGLP LHERFIDIFE QTFSSKKNAK FISMASTSAR RFRWKT CLK IWKEACKLSK TVLDGQQPLP NPQKRQKRLS NQVELRNQWA KFEKEVEDFE KVLTGGSNGF MFSFVEGALV KAVRSGH WV LLDEINLASL ETLEPIGQLL SSYESGILLS ERGDITPITP HKNFRLFGCM NPSTDVGKRE LEPSFRSRFT EIYVHSPD Q NLDDLLSIIQ KYIGSLCIGN EHVIREVAEL YQVAKSLSLD GSLVDGAGQR PHYTVRTLSR TLSYVTEIAP IYGLRRSLY EGFCMSFLTL LDHTSESLLY NHVVRFTLGE LNRDQQNAIL KQIPKVPDHS SYIAFCHYWL RRGSFPVEEQ EHYIITPFVQ KNLLNIARA CSTRMFPILI QGPTSSGKTS MIEYVAKKTG HKFVRINNHE HTDLQEYIGT YVTDDNGSLS FREGVLVEAL R NGYWIVLD ELNLAPTDVL EALNRLLDDN RELFIPETQV LVKPHPEFML FATQNPPGVY AGRKHLSRAF RNRFLEIHFD DI PENELET ILHKRCKIAP SYAAKIVQVF RELSLRRQTT RIFEQKNSFA TLRDLFRWAF REAVGYQQLA ENGYMLLAER ARD QKDKLA VQEVIEKVMK VKIDTDGIYN LDSMEIFQDM SLKEGPLSKV VWTRPMIRLF CLVWRCLLAK EPVLLVGDTG CGKT TVCQI LAECLHKELH IINAHQDTEN GDIIGAQRPV RNRSAVNYSL HSQLCEKFNV QESLDSIDDL IEKFEKLSSS EKNDN LSNL IERQIIKYRS LFEWHDGALV TAMKQGDFFL LDEISLADDS VLERLNSVLE LSRTLTLVEH SNAAVSLTAK DGFAFF ATM NPGGDYGKKE LSPALRNRFT EIWVPPMVDT EDILKIVEGK LHNNKIELAR PLVEYAKWHA NEYLYTDVIS IRDVLSA VE FINACEILDL NLVLFNAVSM VFIDALGSFT TFSLSNNLAS LHAERQRCFA KLNELAGSNI MASKSADISI KFSDSSFF I GDFGIPLGDS VESDSTYSLH TDTTLMNASK VLRALQVLKP ILLEGSPGVG KTSLITALAR ETGHQLVRIN LSDQTDLMD LFGSDVPVEG GEGGQFAWRD APFLAAMRNG HWVLLDELNL ASQSVLEGLN ACLDHRNEAY IPELDKVFKA HPNFRVFAAQ NPQHQGGGR KGLPRSFINR FSVVYVEALK EKDMIEIAAC NYHQVNEDWR LKIIKFMFRL QDNIEKDISF GSFGSPWEFN L RDTLRWLQ LLNDAPKYTC VSPADYLEVM VLHRMRTVED RVRTCELFKE VFDIDYEPRT IGFSLSSQCF KVGHSLLVRD VE RQKTLLD SQNILQSQLP VLESVITCIN KKWPCILVGD TATGKTCILR LLAAIAGAKI KEMAVNSDTD TMDLIGEYEQ IDI SRKASE LFTDLSQQLL NIVIKYRNFD NIFRETSLYT LTTTSFKTHS QAFTLLQKVV DQLDQLKIHE TLVHSLGDIH EKAR KLLAE FSASPAGRFE WFDGYLLKAV EEGHWFVLDN ANLCSPAVLD RLNSLLEHKG VLIVNEKTTE DGHPKTIKPH PNFRL FLTV NPVYGELSRA MRNRGVEIFL LKEALTEIDK KQMSLLEPAP ISSAVDTLAS NISYIKYVFE TMG(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) |
-分子 #2: PHOSPHOAMINOPHOSPHONIC ACID-ADENYLATE ESTER
| 分子 | 名称: PHOSPHOAMINOPHOSPHONIC ACID-ADENYLATE ESTER / タイプ: ligand / ID: 2 / コピー数: 5 / 式: ANP |
|---|---|
| 分子量 | 理論値: 506.196 Da |
| Chemical component information | 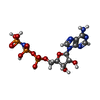 ChemComp-ANP: |
-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | particle |
-試料調製
| 濃度 | 0.15 mg/mL |
|---|---|
| 緩衝液 | pH: 7.5 |
| グリッド | モデル: Quantifoil R1.2/1.3 / 材質: GOLD / メッシュ: 400 / 支持フィルム - 材質: CARBON / 支持フィルム - トポロジー: HOLEY ARRAY / 前処理 - タイプ: GLOW DISCHARGE |
| 凍結 | 凍結剤: ETHANE / チャンバー内湿度: 100 % / チャンバー内温度: 277 K / 装置: FEI VITROBOT MARK IV |
- 電子顕微鏡法
電子顕微鏡法
| 顕微鏡 | FEI TITAN KRIOS |
|---|---|
| 撮影 | フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 検出モード: SUPER-RESOLUTION / デジタル化 - 画像ごとのフレーム数: 1-50 / 実像数: 8446 / 平均露光時間: 15.0 sec. / 平均電子線量: 88.8 e/Å2 |
| 電子線 | 加速電圧: 300 kV / 電子線源:  FIELD EMISSION GUN FIELD EMISSION GUN |
| 電子光学系 | C2レンズ絞り径: 100.0 µm / 照射モード: FLOOD BEAM / 撮影モード: BRIGHT FIELD / Cs: 2.7 mm / 最大 デフォーカス(公称値): 3.0 µm / 最小 デフォーカス(公称値): 1.4 µm / 倍率(公称値): 22500 |
| 試料ステージ | 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER ホルダー冷却材: NITROGEN |
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
+画像解析
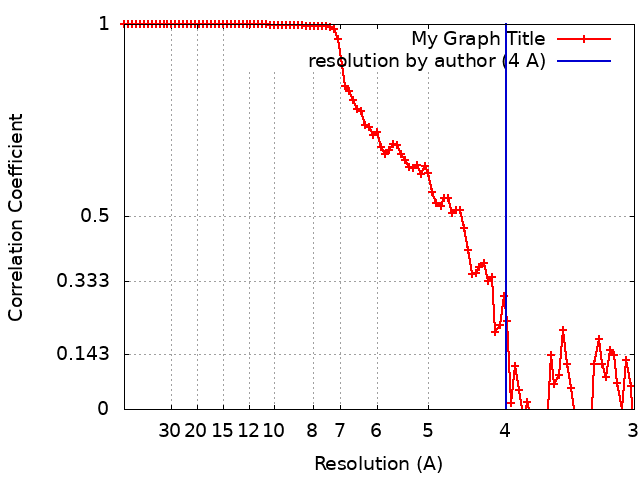
-原子モデル構築 1
| 精密化 | 空間: REAL |
|---|---|
| 得られたモデル |  PDB-6or5: |
