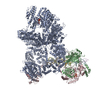
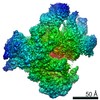
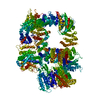
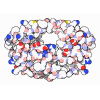
登録情報 データベース : EMDB / ID : EMD-25440タイトル CryoEM structure of DNA-PK complex VIII 複合体 : DNA-PKタンパク質・ペプチド : DNA-dependent protein kinase catalytic subunitタンパク質・ペプチド : X-ray repair cross-complementing protein 5リガンド : MAGNESIUM IONリガンド : ADENOSINE-5'-TRIPHOSPHATE機能・相同性 分子機能 ドメイン・相同性 構成要素
/ / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / 生物種 Human (ヒト) / Homo sapiens (ヒト)手法 / / 解像度 : 3.6 Å Chen X / Liu L / Gellert M / Yang W 資金援助 Organization Grant number 国 National Institutes of Health/National Institute of Diabetes and Digestive and Kidney Disease (NIH/NIDDK)
ジャーナル : Mol Cell / 年 : 2022タイトル : Autophosphorylation transforms DNA-PK from protecting to processing DNA ends.著者 : Lan Liu / Xuemin Chen / Jun Li / Huaibin Wang / Christopher J Buehl / Noah J Goff / Katheryn Meek / Wei Yang / Martin Gellert / 要旨 : The DNA-dependent protein kinase (DNA-PK) initially protects broken DNA ends but then promotes their processing during non-homologous end joining (NHEJ). Before ligation by NHEJ, DNA hairpin ends ... The DNA-dependent protein kinase (DNA-PK) initially protects broken DNA ends but then promotes their processing during non-homologous end joining (NHEJ). Before ligation by NHEJ, DNA hairpin ends generated during V(D)J recombination must be opened by the Artemis nuclease, together with autophosphorylated DNA-PK. Structures of DNA-PK bound to DNA before and after phosphorylation, and in complex with Artemis and a DNA hairpin, reveal an essential functional switch. When bound to open DNA ends in its protection mode, DNA-PK is inhibited for cis-autophosphorylation of the so-called ABCDE cluster but activated for phosphorylation of other targets. In contrast, DNA hairpin ends promote cis-autophosphorylation. Phosphorylation of four Thr residues in ABCDE leads to gross structural rearrangement of DNA-PK, widening the DNA binding groove for Artemis recruitment and hairpin cleavage. Meanwhile, Artemis locks DNA-PK into the kinase-inactive state. Kinase activity and autophosphorylation of DNA-PK are regulated by different DNA ends, feeding forward to coordinate NHEJ events. 履歴 登録 2021年11月16日 - ヘッダ(付随情報) 公開 2022年1月12日 - マップ公開 2022年1月12日 - 更新 2022年1月19日 - 現状 2022年1月19日 処理サイト : RCSB / 状態 : 公開
すべて表示 表示を減らす
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 マップデータ
マップデータ 試料
試料 機能・相同性情報
機能・相同性情報 Human (ヒト) /
Human (ヒト) /  データ登録者
データ登録者 米国, 1件
米国, 1件  引用
引用 構造の表示
構造の表示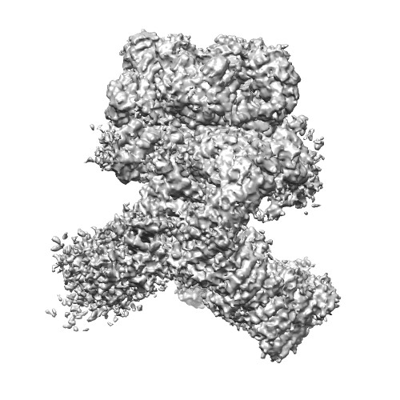
 ダウンロードとリンク
ダウンロードとリンク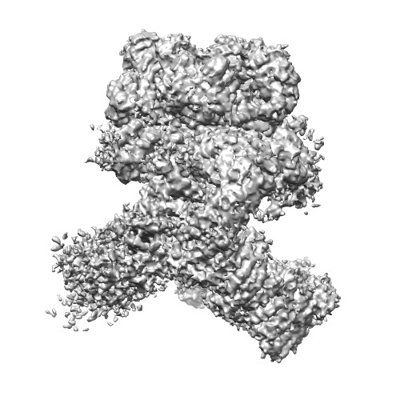 emd_25440.png
emd_25440.png http://ftp.pdbj.org/pub/emdb/structures/EMD-25440
http://ftp.pdbj.org/pub/emdb/structures/EMD-25440





















 試料の構成要素
試料の構成要素
 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN
