 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-8g5z: mRNA decoding in human is kinetically and structurally distinct f... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8g5z | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | mRNA decoding in human is kinetically and structurally distinct from bacteria (GA state) | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | RIBOSOME / Human 80S / tRNA / mRNA eEF1A / eIF5A / tRNA selection | |||||||||
| Function / homology |  Function and homology information Function and homology informationguanyl nucleotide binding / embryonic brain development / translation at presynapse / regulation of translation involved in cellular response to UV / eukaryotic 80S initiation complex / ribosomal protein import into nucleus / regulation of G1 to G0 transition / negative regulation of endoplasmic reticulum unfolded protein response / negative regulation of formation of translation preinitiation complex / kinase activator activity ...guanyl nucleotide binding / embryonic brain development / translation at presynapse / regulation of translation involved in cellular response to UV / eukaryotic 80S initiation complex / ribosomal protein import into nucleus / regulation of G1 to G0 transition / negative regulation of endoplasmic reticulum unfolded protein response / negative regulation of formation of translation preinitiation complex / kinase activator activity / axial mesoderm development / oxidized pyrimidine DNA binding / response to TNF agonist / positive regulation of base-excision repair / positive regulation of intrinsic apoptotic signaling pathway in response to DNA damage by p53 class mediator / protein-DNA complex disassembly / positive regulation of respiratory burst involved in inflammatory response / positive regulation of gastrulation / 90S preribosome assembly / positive regulation of ubiquitin-protein transferase activity / protein tyrosine kinase inhibitor activity / positive regulation of intrinsic apoptotic signaling pathway in response to DNA damage / positive regulation of DNA-templated transcription initiation / IRE1-RACK1-PP2A complex / alpha-beta T cell differentiation / positive regulation of Golgi to plasma membrane protein transport / nucleolus organization / TNFR1-mediated ceramide production / positive regulation of DNA damage response, signal transduction by p53 class mediator / GAIT complex / negative regulation of RNA splicing / neural crest cell differentiation / supercoiled DNA binding / G1 to G0 transition / NF-kappaB complex / negative regulation of DNA repair / cytoplasmic translational initiation / oxidized purine DNA binding / middle ear morphogenesis / cysteine-type endopeptidase activator activity involved in apoptotic process / rRNA modification in the nucleus and cytosol / negative regulation of intrinsic apoptotic signaling pathway in response to hydrogen peroxide / negative regulation of bicellular tight junction assembly / ubiquitin-like protein conjugating enzyme binding / regulation of establishment of cell polarity / negative regulation of phagocytosis / erythrocyte homeostasis / cytoplasmic side of rough endoplasmic reticulum membrane / Formation of the ternary complex, and subsequently, the 43S complex / ion channel inhibitor activity / protein kinase A binding / laminin receptor activity / homeostatic process / pigmentation / Ribosomal scanning and start codon recognition / positive regulation of mitochondrial depolarization / macrophage chemotaxis / Translation initiation complex formation / lung morphogenesis / negative regulation of Wnt signaling pathway / positive regulation of natural killer cell proliferation / fibroblast growth factor binding / Protein hydroxylation / monocyte chemotaxis / BH3 domain binding / negative regulation of translational frameshifting / regulation of adenylate cyclase-activating G protein-coupled receptor signaling pathway / TOR signaling / mTORC1-mediated signalling / SARS-CoV-1 modulates host translation machinery / iron-sulfur cluster binding / positive regulation of GTPase activity / regulation of cell division / cellular response to ethanol / Peptide chain elongation / Selenocysteine synthesis / Formation of a pool of free 40S subunits / negative regulation of protein binding / blastocyst development / Eukaryotic Translation Termination / positive regulation of intrinsic apoptotic signaling pathway by p53 class mediator / protein serine/threonine kinase inhibitor activity / SRP-dependent cotranslational protein targeting to membrane / Response of EIF2AK4 (GCN2) to amino acid deficiency / negative regulation of respiratory burst involved in inflammatory response / ubiquitin ligase inhibitor activity / Viral mRNA Translation / endonucleolytic cleavage to generate mature 3'-end of SSU-rRNA from (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / translation elongation factor activity / protein localization to nucleus / negative regulation of ubiquitin-dependent protein catabolic process / Nonsense Mediated Decay (NMD) independent of the Exon Junction Complex (EJC) / positive regulation of signal transduction by p53 class mediator / GTP hydrolysis and joining of the 60S ribosomal subunit / L13a-mediated translational silencing of Ceruloplasmin expression / Major pathway of rRNA processing in the nucleolus and cytosol / protein targeting / regulation of translational fidelity / positive regulation of microtubule polymerization / phagocytic cup Similarity search - Function | |||||||||
| Biological species |   Homo sapiens (human) Homo sapiens (human) | |||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 2.64 Å | |||||||||
 Authors Authors | Holm, M. / Natchiar, K.S. / Rundlet, E.J. / Myasnikov, A.G. / Altman, R.B. / Blanchard, S.C. | |||||||||
| Funding support | 1items
| |||||||||
 Citation Citation | Journal: Nature / Year: 2023 Title: mRNA decoding in human is kinetically and structurally distinct from bacteria. Authors: Mikael Holm / S Kundhavai Natchiar / Emily J Rundlet / Alexander G Myasnikov / Zoe L Watson / Roger B Altman / Hao-Yuan Wang / Jack Taunton / Scott C Blanchard /   Abstract: In all species, ribosomes synthesize proteins by faithfully decoding messenger RNA (mRNA) nucleotide sequences using aminoacyl-tRNA substrates. Current knowledge of the decoding mechanism derives ...In all species, ribosomes synthesize proteins by faithfully decoding messenger RNA (mRNA) nucleotide sequences using aminoacyl-tRNA substrates. Current knowledge of the decoding mechanism derives principally from studies on bacterial systems. Although key features are conserved across evolution, eukaryotes achieve higher-fidelity mRNA decoding than bacteria. In human, changes in decoding fidelity are linked to ageing and disease and represent a potential point of therapeutic intervention in both viral and cancer treatment. Here we combine single-molecule imaging and cryogenic electron microscopy methods to examine the molecular basis of human ribosome fidelity to reveal that the decoding mechanism is both kinetically and structurally distinct from that of bacteria. Although decoding is globally analogous in both species, the reaction coordinate of aminoacyl-tRNA movement is altered on the human ribosome and the process is an order of magnitude slower. These distinctions arise from eukaryote-specific structural elements in the human ribosome and in the elongation factor eukaryotic elongation factor 1A (eEF1A) that together coordinate faithful tRNA incorporation at each mRNA codon. The distinct nature and timing of conformational changes within the ribosome and eEF1A rationalize how increased decoding fidelity is achieved and potentially regulated in eukaryotic species. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8g5z.cif.gz | 5 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8g5z.ent.gz | Display | PDB format | |
| PDBx/mmJSON format | 8g5z.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/g5/8g5zftp://data.pdbj.org/pub/pdb/validation_reports/g5/8g5z | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  29758MC  8g5yC  8g60C  8g61C  8g6jC  8glpC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |





-Links
 PDBj
PDBj


































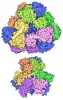








- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
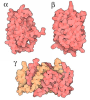
-Components
-RNA chain , 7 types, 7 molecules S2L8L5L7mRAtPt
| #1: RNA chain | Mass: 549676.062 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) |
|---|---|
| #2: RNA chain | Mass: 50171.703 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: GenBank: 555853 |
| #3: RNA chain | Mass: 1195713.500 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) |
| #4: RNA chain | Mass: 38691.914 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: GenBank: 23898 |
| #83: RNA chain | Mass: 3468.098 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) Homo sapiens (human) |
| #84: RNA chain | Mass: 24634.879 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) |
| #85: RNA chain | Mass: 24848.943 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) |
-40S ribosomal protein ... , 19 types, 19 molecules SBSASDSJSCSGSFSHSWSISdSNSRSTSVSYSaScSe
| #5: Protein | Mass: 25859.438 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P61247 |
|---|---|
| #6: Protein | Mass: 24903.479 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P08865 |
| #7: Protein | Mass: 25043.381 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human)References: UniProt: P23396, DNA-(apurinic or apyrimidinic site) lyase |
| #8: Protein | Mass: 21649.633 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P46781 |
| #10: Protein | Mass: 24587.029 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P15880 |
| #11: Protein | Mass: 27471.535 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P62753 |
| #12: Protein | Mass: 21327.723 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P46782 |
| #13: Protein | Mass: 21603.229 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P62081 |
| #14: Protein | Mass: 14734.357 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P62244 |
| #15: Protein | Mass: 24003.012 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P62241 |
| #23: Protein | Mass: 6559.625 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P62273 |
| #24: Protein | Mass: 17128.191 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P62277 |
| #26: Protein | Mass: 15446.961 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P08708 |
| #28: Protein | Mass: 15822.219 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P39019 |
| #29: Protein | Mass: 9166.426 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P63220 |
| #30: Protein | Mass: 15203.022 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P62847 |
| #32: Protein | Mass: 11341.426 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P62854 |
| #34: Protein | Mass: 7263.394 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P62857 |
| #35: Protein | Mass: 6668.938 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P62861 |
+Protein , 25 types, 25 molecules SESQSUSKSOSXSMSSSLSPSZSbSfSgLzLALELqLXLWLbLmLo5AEF
+60S ribosomal protein ... , 36 types, 36 molecules LBLCLJLHLGLKLOLLLVLMLaLNLILDLQLRLSLTLPLULYLZLrLhLFLcLdLeLfLg...
-Non-polymers , 12 types, 875 molecules 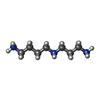






| #88: Chemical | ChemComp-SPD /  Mass: 145.246 Da / Num. of mol.: 10 / Source method: obtained synthetically / Formula: C7H19N3 Mass: 145.246 Da / Num. of mol.: 10 / Source method: obtained synthetically / Formula: C7H19N3#89: Chemical | ChemComp-K /  Mass: 39.098 Da / Num. of mol.: 45 / Source method: obtained synthetically / Formula: K Mass: 39.098 Da / Num. of mol.: 45 / Source method: obtained synthetically / Formula: K#90: Chemical | ChemComp-MG /  Mass: 24.305 Da / Num. of mol.: 433 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 433 / Source method: obtained synthetically / Formula: Mg#91: Chemical | ChemComp-ANM / |  Mass: 265.305 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C14H19NO4 / Feature type: SUBJECT OF INVESTIGATION Mass: 265.305 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C14H19NO4 / Feature type: SUBJECT OF INVESTIGATION#92: Chemical | ChemComp-PUT /  Mass: 88.151 Da / Num. of mol.: 8 / Source method: obtained synthetically / Formula: C4H12N2 Mass: 88.151 Da / Num. of mol.: 8 / Source method: obtained synthetically / Formula: C4H12N2#93: Chemical | ChemComp-3H3 / |  Mass: 457.559 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C26H35NO6 / Feature type: SUBJECT OF INVESTIGATION Mass: 457.559 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C26H35NO6 / Feature type: SUBJECT OF INVESTIGATION#94: Chemical | ChemComp-ZN /  Mass: 65.409 Da / Num. of mol.: 8 / Source method: obtained synthetically / Formula: Zn Mass: 65.409 Da / Num. of mol.: 8 / Source method: obtained synthetically / Formula: Zn#95: Chemical | ChemComp-PHE / |  Type: L-peptide linking / Mass: 165.189 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C9H11NO2 Type: L-peptide linking / Mass: 165.189 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C9H11NO2#96: Chemical | ChemComp-MET / |  Type: L-peptide linking / Mass: 149.211 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C5H11NO2S Type: L-peptide linking / Mass: 149.211 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C5H11NO2S#97: Chemical | ChemComp-GSP / |  Mass: 539.246 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H16N5O13P3S Mass: 539.246 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H16N5O13P3S#98: Chemical | ChemComp-ZIY / |  Mass: 1110.339 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C57H87N7O15 / Feature type: SUBJECT OF INVESTIGATION Mass: 1110.339 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C57H87N7O15 / Feature type: SUBJECT OF INVESTIGATION#99: Water | ChemComp-HOH / | Mass: 18.015 Da / Num. of mol.: 365 / Source method: isolated from a natural source / Formula: H2O |
|---|
-Details
| Has ligand of interest | Y |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: Human ribosome 80S with P-site tRNA, A-site tRNA eEF1A and eIF5A1 Type: RIBOSOME / Entity ID: #1-#48, #50-#83, #86-#87 / Source: MULTIPLE SOURCES |
|---|---|
| Molecular weight | Value: 5.5 MDa / Experimental value: YES |
| Buffer solution | pH: 7.5 |
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Specimen support | Grid material: GOLD / Grid mesh size: 300 divisions/in. / Grid type: Quantifoil R1.2/1.3 |
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal defocus max: 1500 nm / Nominal defocus min: 500 nm |
| Image recording | Electron dose: 79 e/Å2 / Film or detector model: GATAN K3 (6k x 4k) |
- Processing
Processing
| EM software | Name: PHENIX / Version: 1.19_4092: / Category: model refinement | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING ONLY | ||||||||||||||||||||||||
| 3D reconstruction | Resolution: 2.64 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 20290 / Symmetry type: POINT | ||||||||||||||||||||||||
| Refine LS restraints |
|
