 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 7nfe | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | Cryo-EM structure of NHEJ super-complex (monomer) | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | DNA BINDING PROTEIN / NHEJ / DNA-PKcs / Ku70/80 / XLF / XRCC4 / DNA-LigaseIV | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報FHA domain binding / positive regulation of chromosome organization / positive regulation of ligase activity / DNA ligase IV complex / positive regulation of platelet formation / DNA ligase activity / Ku70:Ku80 complex / DN2 thymocyte differentiation / negative regulation of t-circle formation / DNA ligase (ATP) ...FHA domain binding / positive regulation of chromosome organization / positive regulation of ligase activity / DNA ligase IV complex / positive regulation of platelet formation / DNA ligase activity / Ku70:Ku80 complex / DN2 thymocyte differentiation / negative regulation of t-circle formation / DNA ligase (ATP) / T cell receptor V(D)J recombination / DNA end binding / pro-B cell differentiation / small-subunit processome assembly / positive regulation of lymphocyte differentiation / DNA ligase (ATP) activity / DNA-dependent protein kinase activity / DNA-dependent protein kinase complex / histone H2AXS139 kinase activity / DNA-dependent protein kinase-DNA ligase 4 complex / nonhomologous end joining complex / immunoglobulin V(D)J recombination / nucleotide-excision repair, DNA gap filling / immature B cell differentiation / single strand break repair / regulation of smooth muscle cell proliferation / positive regulation of double-strand break repair via nonhomologous end joining / cellular response to X-ray / V(D)J recombination / nuclear telomere cap complex / double-strand break repair via alternative nonhomologous end joining / double-strand break repair via classical nonhomologous end joining / protein localization to site of double-strand break / regulation of epithelial cell proliferation / telomere capping / isotype switching / Cytosolic sensors of pathogen-associated DNA / IRF3-mediated induction of type I IFN / positive regulation of neurogenesis / regulation of hematopoietic stem cell differentiation / regulation of telomere maintenance / recombinational repair / U3 snoRNA binding / protein localization to chromosome, telomeric region / DNA biosynthetic process / maturation of 5.8S rRNA / T cell lineage commitment / response to ionizing radiation / cellular response to lithium ion / cellular hyperosmotic salinity response / negative regulation of cGAS/STING signaling pathway / 2-LTR circle formation / telomeric DNA binding / B cell lineage commitment / hematopoietic stem cell proliferation / ligase activity / negative regulation of protein phosphorylation / positive regulation of protein kinase activity / site of DNA damage / 付加脱離酵素(リアーゼ); 炭素-酸素リアーゼ類; その他の炭素-酸素リアーゼ / T cell differentiation / peptidyl-threonine phosphorylation / somatic stem cell population maintenance / 5'-deoxyribose-5-phosphate lyase activity / response to X-ray / hematopoietic stem cell differentiation / ectopic germ cell programmed cell death / ATP-dependent activity, acting on DNA / chromosome organization / telomere maintenance via telomerase / somitogenesis / SUMOylation of DNA damage response and repair proteins / condensed chromosome / DNA polymerase binding / neurogenesis / mitotic G1 DNA damage checkpoint signaling / cellular response to ionizing radiation / telomere maintenance / activation of innate immune response / DNA helicase activity / cyclin binding / positive regulation of erythrocyte differentiation / positive regulation of translation / cellular response to leukemia inhibitory factor / B cell differentiation / central nervous system development / stem cell proliferation / response to gamma radiation / small-subunit processome / Nonhomologous End-Joining (NHEJ) / enzyme activator activity / cellular response to gamma radiation / regulation of circadian rhythm / protein destabilization / protein-DNA complex / base-excision repair / brain development / peptidyl-serine phosphorylation / protein modification process / double-strand break repair via nonhomologous end joining 類似検索 - 分子機能 | ||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 4.29 Å | ||||||
 データ登録者 データ登録者 | Chaplin, A.K. / Hardwick, S.W. / Kefala Stavridi, A. / Chirgadze, D.Y. / Blundell, T.L. | ||||||
| 資金援助 |  英国, 1件 英国, 1件
| ||||||
 引用 引用 | ジャーナル: Mol Cell / 年: 2021 タイトル: Cryo-EM of NHEJ supercomplexes provides insights into DNA repair. 著者: Amanda K Chaplin / Steven W Hardwick / Antonia Kefala Stavridi / Christopher J Buehl / Noah J Goff / Virginie Ropars / Shikang Liang / Taiana Maia De Oliveira / Dimitri Y Chirgadze / Katheryn ...著者: Amanda K Chaplin / Steven W Hardwick / Antonia Kefala Stavridi / Christopher J Buehl / Noah J Goff / Virginie Ropars / Shikang Liang / Taiana Maia De Oliveira / Dimitri Y Chirgadze / Katheryn Meek / Jean-Baptiste Charbonnier / Tom L Blundell /   要旨: Non-homologous end joining (NHEJ) is one of two critical mechanisms utilized in humans to repair DNA double-strand breaks (DSBs). Unrepaired or incorrect repair of DSBs can lead to apoptosis or ...Non-homologous end joining (NHEJ) is one of two critical mechanisms utilized in humans to repair DNA double-strand breaks (DSBs). Unrepaired or incorrect repair of DSBs can lead to apoptosis or cancer. NHEJ involves several proteins, including the Ku70/80 heterodimer, DNA-dependent protein kinase catalytic subunit (DNA-PKcs), X-ray cross-complementing protein 4 (XRCC4), XRCC4-like factor (XLF), and ligase IV. These core proteins bind DSBs and ligate the damaged DNA ends. However, details of the structural assembly of these proteins remain unclear. Here, we present cryo-EM structures of NHEJ supercomplexes that are composed of these core proteins and DNA, revealing the detailed structural architecture of this assembly. We describe monomeric and dimeric forms of this supercomplex and also propose the existence of alternate dimeric forms of long-range synaptic complexes. Finally, we show that mutational disruption of several structural features within these NHEJ complexes negatively affects DNA repair. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7nfe.cif.gz | 1 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7nfe.ent.gz | 808.7 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 7nfe.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 7nfe_validation.pdf.gz | 1.6 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 7nfe_full_validation.pdf.gz | 1.7 MB | 表示 | |
| XML形式データ | 7nfe_validation.xml.gz | 160.4 KB | 表示 | |
| CIF形式データ | 7nfe_validation.cif.gz | 244.5 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/nf/7nfeftp://data.pdbj.org/pub/pdb/validation_reports/nf/7nfe | HTTPS FTP |
-関連構造データ

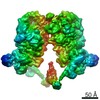


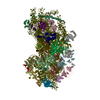

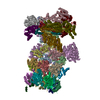
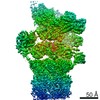
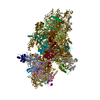
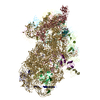
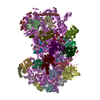
-リンク
 PDBj
PDBj












































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 4種, 6分子 AFGHIJ
| #1: タンパク質 | 分子量: 472056.281 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 細胞株: HELA参照: UniProt: P78527, non-specific serine/threonine protein kinase | ||||
|---|---|---|---|---|---|
| #4: タンパク質 | 分子量: 33372.234 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: NHEJ1, XLF / 発現宿主:  #5: タンパク質 | 分子量: 38337.703 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: XRCC4 / 発現宿主: #6: タンパク質 | | 分子量: 104124.953 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: LIG4 / 発現宿主: |
-X-ray repair cross-complementing protein ... , 2種, 2分子 BC
| #2: タンパク質 | 分子量: 69945.039 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: XRCC6, G22P1発現宿主: Insect cell expression vector pTIE1 (その他) 参照: UniProt: P12956, 加水分解酵素; 酸無水物に作用; 酸無水物に作用・細胞または細胞小器官の運動に関与, 付加脱離酵素(リアーゼ); 炭素- ...参照: UniProt: P12956, 加水分解酵素; 酸無水物に作用; 酸無水物に作用・細胞または細胞小器官の運動に関与, 付加脱離酵素(リアーゼ); 炭素-酸素リアーゼ類; その他の炭素-酸素リアーゼ |
|---|---|
| #3: タンパク質 | 分子量: 82812.438 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: XRCC5, G22P2発現宿主: Insect cell expression vector pTIE1 (その他) 参照: UniProt: P13010, 加水分解酵素; 酸無水物に作用; 酸無水物に作用・細胞または細胞小器官の運動に関与 |
-DNA鎖 , 2種, 2分子 DE
| #7: DNA鎖 | 分子量: 7367.843 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) |
|---|---|
| #8: DNA鎖 | 分子量: 7402.821 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 値: 0.8 MDa / 実験値: NO | ||||||||||||||||||||||||||||||||||||
| 由来(天然) |
| ||||||||||||||||||||||||||||||||||||
| 由来(組換発現) |
| ||||||||||||||||||||||||||||||||||||
| 緩衝液 | pH: 7.4 | ||||||||||||||||||||||||||||||||||||
| 試料 | 濃度: 3 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||||||||
| 急速凍結 | 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 130000 X / Cs: 2.7 mm / C2レンズ絞り径: 50 µm |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 平均露光時間: 1.3 sec. / 電子線照射量: 46.8 e/Å2 フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) |
- 解析
解析
| ソフトウェア |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 749185 | ||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||
| 3次元再構成 | 解像度: 4.29 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 45943 / 対称性のタイプ: POINT | ||||||||||||||||||||||||
| 精密化 | 交差検証法: NONE 立体化学のターゲット値: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 358.24 Å2 | ||||||||||||||||||||||||
| 拘束条件 |
|
