ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 7aoa | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
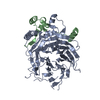
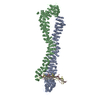
| タイトル | Structure of the extended MTA1/HDAC1/MBD2/RBBP4 NURD deacetylase complex | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | TRANSCRIPTION / Deacetylase / Complex | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報Loss of MECP2 binding ability to 5mC-DNA / Krueppel-associated box domain binding / Repression of WNT target genes / MECP2 regulates transcription of neuronal ligands / satellite DNA binding / CAF-1 complex / p75NTR negatively regulates cell cycle via SC1 / epidermal cell differentiation / histone decrotonylase activity / ventricular cardiac muscle tissue development ...Loss of MECP2 binding ability to 5mC-DNA / Krueppel-associated box domain binding / Repression of WNT target genes / MECP2 regulates transcription of neuronal ligands / satellite DNA binding / CAF-1 complex / p75NTR negatively regulates cell cycle via SC1 / epidermal cell differentiation / histone decrotonylase activity / ventricular cardiac muscle tissue development / fungiform papilla formation / negative regulation of androgen receptor signaling pathway / NURF complex / regulation of cell fate specification / negative regulation of stem cell population maintenance / endoderm development / DNA replication-dependent chromatin assembly / maternal behavior / Transcription of E2F targets under negative control by p107 (RBL1) and p130 (RBL2) in complex with HDAC1 / regulation of stem cell differentiation / NuRD complex / DNA methylation-dependent heterochromatin formation / siRNA binding / protein deacetylation / ESC/E(Z) complex / Transcription of E2F targets under negative control by DREAM complex / STAT3 nuclear events downstream of ALK signaling / Polo-like kinase mediated events / positive regulation of protein autoubiquitination / histone deacetylase / C2H2 zinc finger domain binding / methyl-CpG binding / protein lysine deacetylase activity / positive regulation of signaling receptor activity / 加水分解酵素; ペプチド以外のCN結合加水分解酵素; 鎖状アミドに作用 / embryonic digit morphogenesis / negative regulation of gene expression, epigenetic / positive regulation of oligodendrocyte differentiation / histone deacetylase activity / response to ionizing radiation / Sin3-type complex / positive regulation of stem cell population maintenance / G1/S-Specific Transcription / ATPase complex / cellular response to platelet-derived growth factor stimulus / Notch-HLH transcription pathway / eyelid development in camera-type eye / oligodendrocyte differentiation / Transcriptional Regulation by E2F6 / E-box binding / entrainment of circadian clock by photoperiod / odontogenesis of dentin-containing tooth / locomotor rhythm / RNA Polymerase I Transcription Initiation / SUMOylation of transcription factors / histone deacetylase complex / hair follicle placode formation / Regulation of MECP2 expression and activity / cellular response to organic cyclic compound / G0 and Early G1 / NF-kappaB binding / positive regulation of Wnt signaling pathway / negative regulation by host of viral transcription / RNA polymerase II core promoter sequence-specific DNA binding / embryonic organ development / FOXO-mediated transcription of oxidative stress, metabolic and neuronal genes / Nuclear events stimulated by ALK signaling in cancer / heterochromatin / Cyclin E associated events during G1/S transition / response to mechanical stimulus / MECP2 regulates neuronal receptors and channels / core promoter sequence-specific DNA binding / negative regulation of intrinsic apoptotic signaling pathway / Cyclin A:Cdk2-associated events at S phase entry / negative regulation of canonical NF-kappaB signal transduction / nucleosome binding / Deposition of new CENPA-containing nucleosomes at the centromere / Regulation of TP53 Activity through Acetylation / transcription repressor complex / RNA Polymerase I Promoter Opening / SUMOylation of chromatin organization proteins / negative regulation of cell migration / response to nutrient levels / transcription corepressor binding / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / PRC2 methylates histones and DNA / Regulation of PTEN gene transcription / Defective pyroptosis / Deactivation of the beta-catenin transactivating complex / HDACs deacetylate histones / hippocampus development / promoter-specific chromatin binding / Downregulation of SMAD2/3:SMAD4 transcriptional activity / SMAD2/SMAD3:SMAD4 heterotrimer regulates transcription / positive regulation of smooth muscle cell proliferation / negative regulation of transforming growth factor beta receptor signaling pathway / Formation of the beta-catenin:TCF transactivating complex / circadian regulation of gene expression / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / NoRC negatively regulates rRNA expression 類似検索 - 分子機能 | ||||||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 19.4 Å | ||||||||||||
 データ登録者 データ登録者 | Millard, C.J. / Fairall, L. / Ragan, T.J. / Savva, C.G. / Schwabe, J.W.R. | ||||||||||||
| 資金援助 |  英国, 3件 英国, 3件
| ||||||||||||
 引用 引用 | ジャーナル: Nucleic Acids Res / 年: 2020 タイトル: The topology of chromatin-binding domains in the NuRD deacetylase complex. 著者: Christopher J Millard / Louise Fairall / Timothy J Ragan / Christos G Savva / John W R Schwabe / 要旨: Class I histone deacetylase complexes play essential roles in many nuclear processes. Whilst they contain a common catalytic subunit, they have diverse modes of action determined by associated ...Class I histone deacetylase complexes play essential roles in many nuclear processes. Whilst they contain a common catalytic subunit, they have diverse modes of action determined by associated factors in the distinct complexes. The deacetylase module from the NuRD complex contains three protein domains that control the recruitment of chromatin to the deacetylase enzyme, HDAC1/2. Using biochemical approaches and cryo-electron microscopy, we have determined how three chromatin-binding domains (MTA1-BAH, MBD2/3 and RBBP4/7) are assembled in relation to the core complex so as to facilitate interaction of the complex with the genome. We observe a striking arrangement of the BAH domains suggesting a potential mechanism for binding to di-nucleosomes. We also find that the WD40 domains from RBBP4 are linked to the core with surprising flexibility that is likely important for chromatin engagement. A single MBD2 protein binds asymmetrically to the dimerisation interface of the complex. This symmetry mismatch explains the stoichiometry of the complex. Finally, our structures suggest how the holo-NuRD might assemble on a di-nucleosome substrate. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7aoa.cif.gz | 466.6 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7aoa.ent.gz | 365.1 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 7aoa.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 7aoa_validation.pdf.gz | 1.2 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 7aoa_full_validation.pdf.gz | 1.3 MB | 表示 | |
| XML形式データ | 7aoa_validation.xml.gz | 64.5 KB | 表示 | |
| CIF形式データ | 7aoa_validation.cif.gz | 102.4 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/ao/7aoaftp://data.pdbj.org/pub/pdb/validation_reports/ao/7aoa | HTTPS FTP |
-関連構造データ







-リンク
 PDBj
PDBj



























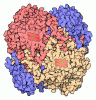
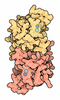







- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 4種, 7分子 CDAEBFG
| #1: タンパク質 | 分子量: 43323.625 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: MBD2 / 細胞株 (発現宿主): HEK293 / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: Q9UBB5 | ||||
|---|---|---|---|---|---|
| #2: タンパク質 | 分子量: 80904.312 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: MTA1 / 細胞株 (発現宿主): HEK293 / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: Q13330#3: タンパク質 | 分子量: 55178.906 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: HDAC1, RPD3L1 / 細胞株 (発現宿主): HEK293 / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: Q13547, histone deacetylase#4: タンパク質 | 分子量: 47709.527 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: RBBP4, RBAP48 / 細胞株 (発現宿主): HEK293 / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: Q09028 |
-非ポリマー , 3種, 8分子 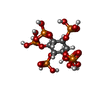


| #5: 化合物 |  分子量: 660.035 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C6H18O24P6 分子量: 660.035 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C6H18O24P6#6: 化合物 |  分子量: 65.409 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Zn#7: 化合物 | ChemComp-K /  分子量: 39.098 Da / 分子数: 4 / 由来タイプ: 合成 / 式: K 分子量: 39.098 Da / 分子数: 4 / 由来タイプ: 合成 / 式: K |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | N |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Extended NuRD deacetylase complex containing two copies of MTA1, HDAC1 and RBBP4 and a single copy of MBD2 タイプ: COMPLEX / Entity ID: #1-#4 / 由来: RECOMBINANT | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 値: 0.34 MDa / 実験値: NO | ||||||||||||||||||||||||
| 由来(天然) | 生物種: Homo sapiens (ヒト) | ||||||||||||||||||||||||
| 由来(組換発現) | 生物種: Homo sapiens (ヒト) | ||||||||||||||||||||||||
| 緩衝液 | pH: 7.5 | ||||||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||||||
| 試料 | 濃度: 0.1 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||
| 試料支持 | 詳細: 40 mA for 120 sec / グリッドの材料: GOLD / グリッドのサイズ: 300 divisions/in. / グリッドのタイプ: Quantifoil R1.2/1.3 | ||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277 K / 詳細: Blot for 3 seconds, blot force 10 |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 75000 X / 倍率(補正後): 129629 X / 最小 デフォーカス(公称値): 500 nm / Calibrated defocus min: 500 nm / Cs: 2.7 mm / C2レンズ絞り径: 50 µm / アライメント法: COMA FREE |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER 最低温度: 100 K |
| 撮影 | 平均露光時間: 60 sec. / 電子線照射量: 34 e/Å2 / 検出モード: COUNTING フィルム・検出器のモデル: FEI FALCON III (4k x 4k) 撮影したグリッド数: 1 / 実像数: 1902 |
| 電子光学装置 | 位相板: VOLTA PHASE PLATE |
| 画像スキャン | サンプリングサイズ: 14 µm / 横: 4096 / 縦: 4096 |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 55799 | ||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 19.4 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 10066 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT | ||||||||||||||||||||||||||||||||||||
| 原子モデル構築 |
|