 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 5kgf | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Structural model of 53BP1 bound to a ubiquitylated and methylated nucleosome, at 4.5 A resolution | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | STRUCTURAL PROTEIN/DNA / DNA / chromatin / 53BP1 / STRUCTURAL PROTEIN-DNA complex | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報ubiquitin-modified histone reader activity / positive regulation of isotype switching / histone H4K20me2 reader activity / cellular response to X-ray / double-strand break repair via classical nonhomologous end joining / protein localization to site of double-strand break / DNA repair complex / positive regulation of intrinsic apoptotic signaling pathway by p53 class mediator / symbiont entry into host cell via disruption of host cell glycocalyx / symbiont entry into host cell via disruption of host cell envelope ...ubiquitin-modified histone reader activity / positive regulation of isotype switching / histone H4K20me2 reader activity / cellular response to X-ray / double-strand break repair via classical nonhomologous end joining / protein localization to site of double-strand break / DNA repair complex / positive regulation of intrinsic apoptotic signaling pathway by p53 class mediator / symbiont entry into host cell via disruption of host cell glycocalyx / symbiont entry into host cell via disruption of host cell envelope / telomeric DNA binding / virus tail / : / histone reader activity / SUMOylation of transcription factors / negative regulation of double-strand break repair via homologous recombination / Replacement of protamines by nucleosomes in the male pronucleus / Packaging Of Telomere Ends / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / Deposition of new CENPA-containing nucleosomes at the centromere / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / Inhibition of DNA recombination at telomere / RNA Polymerase I Promoter Opening / Meiotic synapsis / Assembly of the ORC complex at the origin of replication / Regulation of endogenous retroelements by the Human Silencing Hub (HUSH) complex / DNA damage checkpoint signaling / DNA methylation / Condensation of Prophase Chromosomes / Chromatin modifications during the maternal to zygotic transition (MZT) / SIRT1 negatively regulates rRNA expression / HCMV Late Events / replication fork / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / innate immune response in mucosa / PRC2 methylates histones and DNA / Regulation of endogenous retroelements by KRAB-ZFP proteins / Defective pyroptosis / Regulation of endogenous retroelements by Piwi-interacting RNAs (piRNAs) / transcription coregulator activity / HDACs deacetylate histones / Nonhomologous End-Joining (NHEJ) / RNA Polymerase I Promoter Escape / Transcriptional regulation by small RNAs / Formation of the beta-catenin:TCF transactivating complex / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / G2/M DNA damage checkpoint / Metalloprotease DUBs / NoRC negatively regulates rRNA expression / protein homooligomerization / kinetochore / B-WICH complex positively regulates rRNA expression / DNA Damage/Telomere Stress Induced Senescence / Meiotic recombination / double-strand break repair via nonhomologous end joining / Pre-NOTCH Transcription and Translation / RMTs methylate histone arginines / Activation of anterior HOX genes in hindbrain development during early embryogenesis / Transcriptional regulation of granulopoiesis / UCH proteinases / HCMV Early Events / antimicrobial humoral immune response mediated by antimicrobial peptide / structural constituent of chromatin / p53 binding / antibacterial humoral response / E3 ubiquitin ligases ubiquitinate target proteins / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / nucleosome / site of double-strand break / heterochromatin formation / RUNX1 regulates transcription of genes involved in differentiation of HSCs / nucleosome assembly / Processing of DNA double-strand break ends / HATs acetylate histones / Senescence-Associated Secretory Phenotype (SASP) / MLL4 and MLL3 complexes regulate expression of PPARG target genes in adipogenesis and hepatic steatosis / Oxidative Stress Induced Senescence / histone binding / damaged DNA binding / Estrogen-dependent gene expression / RNA polymerase II-specific DNA-binding transcription factor binding / transcription coactivator activity / chromosome, telomeric region / Ub-specific processing proteases / defense response to Gram-positive bacterium / nuclear body / Amyloid fiber formation / protein heterodimerization activity / DNA damage response / positive regulation of DNA-templated transcription / enzyme binding / positive regulation of transcription by RNA polymerase II / extracellular space / DNA binding / extracellular exosome / nucleoplasm / identical protein binding 類似検索 - 分子機能 | ||||||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト)synthetic construct (人工物) | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 4.54 Å | ||||||||||||
 データ登録者 データ登録者 | Wilson, M.D. / Benlekbir, S. / Sicheri, F. / Rubinstein, J.L. / Durocher, D. | ||||||||||||
| 資金援助 |  カナダ, 3件 カナダ, 3件
| ||||||||||||
 引用 引用 | ジャーナル: Nature / 年: 2016 タイトル: The structural basis of modified nucleosome recognition by 53BP1. 要旨: DNA double-strand breaks (DSBs) elicit a histone modification cascade that controls DNA repair. This pathway involves the sequential ubiquitination of histones H1 and H2A by the E3 ubiquitin ligases ...DNA double-strand breaks (DSBs) elicit a histone modification cascade that controls DNA repair. This pathway involves the sequential ubiquitination of histones H1 and H2A by the E3 ubiquitin ligases RNF8 and RNF168, respectively. RNF168 ubiquitinates H2A on lysine 13 and lysine 15 (refs 7, 8) (yielding H2AK13ub and H2AK15ub, respectively), an event that triggers the recruitment of 53BP1 (also known as TP53BP1) to chromatin flanking DSBs. 53BP1 binds specifically to H2AK15ub-containing nucleosomes through a peptide segment termed the ubiquitination-dependent recruitment motif (UDR), which requires the simultaneous engagement of histone H4 lysine 20 dimethylation (H4K20me2) by its tandem Tudor domain. How 53BP1 interacts with these two histone marks in the nucleosomal context, how it recognizes ubiquitin, and how it discriminates between H2AK13ub and H2AK15ub is unknown. Here we present the electron cryomicroscopy (cryo-EM) structure of a dimerized human 53BP1 fragment bound to a H4K20me2-containing and H2AK15ub-containing nucleosome core particle (NCP-ubme) at 4.5 Å resolution. The structure reveals that H4K20me2 and H2AK15ub recognition involves intimate contacts with multiple nucleosomal elements including the acidic patch. Ubiquitin recognition by 53BP1 is unusual and involves the sandwiching of the UDR segment between ubiquitin and the NCP surface. The selectivity for H2AK15ub is imparted by two arginine fingers in the H2A amino-terminal tail, which straddle the nucleosomal DNA and serve to position ubiquitin over the NCP-bound UDR segment. The structure of the complex between NCP-ubme and 53BP1 reveals the basis of 53BP1 recruitment to DSB sites and illuminates how combinations of histone marks and nucleosomal elements cooperate to produce highly specific chromatin responses, such as those elicited following chromosome breaks. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 5kgf.cif.gz | 365.4 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb5kgf.ent.gz | 274.6 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 5kgf.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 5kgf_validation.pdf.gz | 1 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 5kgf_full_validation.pdf.gz | 1.1 MB | 表示 | |
| XML形式データ | 5kgf_validation.xml.gz | 47.5 KB | 表示 | |
| CIF形式データ | 5kgf_validation.cif.gz | 72.3 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/kg/5kgfftp://data.pdbj.org/pub/pdb/validation_reports/kg/5kgf | HTTPS FTP |
-関連構造データ

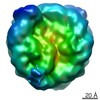
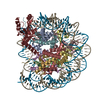
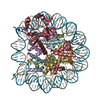
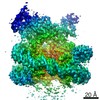

-リンク
 PDBj
PDBj












































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 5種, 10分子 AEBFCGDHOM
| #1: タンパク質 | 分子量: 15421.101 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) 発現宿主:  #2: タンパク質 | 分子量: 11439.511 Da / 分子数: 2 / Mutation: K20C / 由来タイプ: 組換発現 / 詳細: cysteine alkylation at position 20 由来: (組換発現) 詳細 (発現宿主): pDD2349 / 発現宿主: #3: タンパク質 | 分子量: 14163.539 Da / 分子数: 2 / Mutation: K13R, T16S, K36R / 由来タイプ: 組換発現 詳細: Isopeptide amide crosslink between K15 of H2A and G76 of ubiquitin 由来: (組換発現) Homo sapiens (ヒト)遺伝子: HIST1H2AG, H2AFP, HIST1H2AI, H2AFC, HIST1H2AK, H2AFD, HIST1H2AL, H2AFI, HIST1H2AM, H2AFN 発現宿主: #4: タンパク質 | 分子量: 13937.213 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: HIST1H2BC, H2BFL, HIST1H2BE, H2BFH, HIST1H2BF, H2BFG, HIST1H2BG, H2BFA, HIST1H2BI, H2BFK 発現宿主: #8: タンパク質 | 分子量: 8576.831 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: UBB / 発現宿主: |
|---|
-DNA鎖 , 2種, 2分子 IJ
| #5: DNA鎖 | 分子量: 44520.383 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) synthetic construct (人工物) / 発現宿主: |
|---|---|
| #6: DNA鎖 | 分子量: 44991.660 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) synthetic construct (人工物) / 発現宿主: |
-タンパク質・ペプチド , 1種, 2分子 LK
| #7: タンパク質・ペプチド | 分子量: 2376.757 Da / 分子数: 2 / 由来タイプ: 組換発現 / 詳細: full protein not modeled / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: TP53BP1 / 発現宿主: |
|---|
-詳細
| Has protein modification | Y |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 由来(天然) |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 由来(組換発現) |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 緩衝液 | pH: 7.5 詳細: High concentration NCP-ubme/GST-53BP1 complex at 200 mM salt was diluted just prior to grid freezing. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 緩衝液成分 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 試料 | 濃度: 0.6 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 試料支持 | グリッドの材料: COPPER/RHODIUM / グリッドのサイズ: 400 divisions/in. / グリッドのタイプ: electron micrsocopy sciences | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK III / 凍結剤: ETHANE-PROPANE / 湿度: 100 % / 凍結前の試料温度: 277 K 詳細: Plunged into liquid ethane-propane (FEI VITROBOT MARK III) |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Tecnai F20 / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TECNAI F20 |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 200 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 200 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 25000 X / 倍率(補正後): 34483 X / Cs: 2 mm / C2レンズ絞り径: 30 µm / アライメント法: COMA FREE |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: GATAN 626 SINGLE TILT LIQUID NITROGEN CRYO TRANSFER HOLDER |
| 撮影 | 平均露光時間: 0.5 sec. / 電子線照射量: 36 e/Å2 / 検出モード: COUNTING フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 撮影したグリッド数: 2 / 実像数: 319 |
| 画像スキャン | 動画フレーム数/画像: 30 / 利用したフレーム数/画像: 1-30 |
- 解析
解析
| ソフトウェア | 名称: PHENIX / バージョン: 1.10.1_2155: / 分類: 精密化 | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 174185 詳細: Automatically picked from roughly 3000 particles using a manually picked template | ||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C2 (2回回転対称) | ||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 4.54 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 45361 / アルゴリズム: FOURIER SPACE / クラス平均像の数: 9 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | B value: 207.5 / プロトコル: RIGID BODY FIT / 空間: REAL 詳細: The atomic models of Widom-601 DNA (PDB ID 3LZ0), octameric histones (PDB ID 1KX5), ubiquitin (PDB ID 1UBI), and H4K20me2/53BP1 tandem Tudor domain (PDB ID 2IG0) were fitted without allowing ...詳細: The atomic models of Widom-601 DNA (PDB ID 3LZ0), octameric histones (PDB ID 1KX5), ubiquitin (PDB ID 1UBI), and H4K20me2/53BP1 tandem Tudor domain (PDB ID 2IG0) were fitted without allowing flexibility into the 3D maps using UCSF Chimera. Segmentation was performed in UCSF Chimera. For the NCP-ubme structure the ubiquitin segmentation was further modified to remove obvious NCP density from the ubiquitin segment. The H2A/H2B sequence was mutated to the human H2AK13R/K36R and H2B manually in UCSF Chimera. A polyalanine model of the UDR was built within the UDR density in Coot, which compared well to predicted structures generated by Rosetta. The UDR model was mutated and fitted using UCSF Chimera, followed by iterative rounds of real-space refinement in PHENIX and model optimization in Coot. All figures were prepared in UCSF Chimera. | ||||||||||||||||||||||||||||||||||||
| 精密化 | 最高解像度: 4.54 Å | ||||||||||||||||||||||||||||||||||||
| 拘束条件 |
|
