 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: EMDB / ID: EMD-20849 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
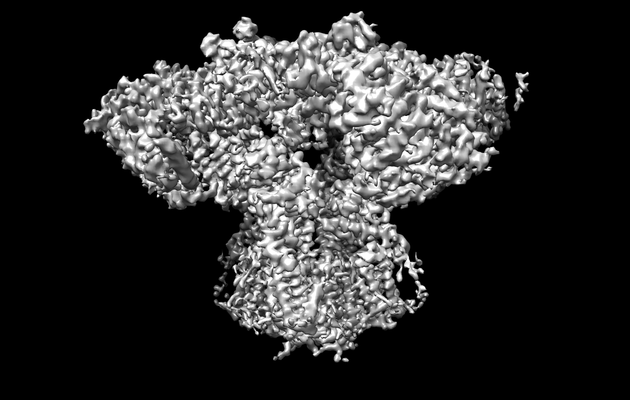
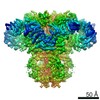
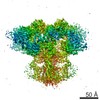
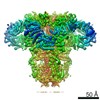
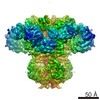
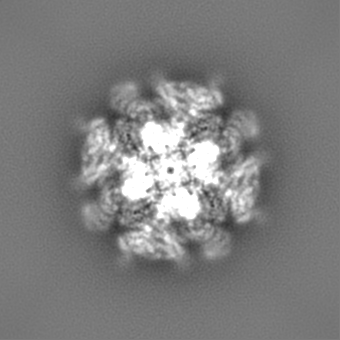
| タイトル | Cryo-EM structure of type 3 IP3 receptor revealing presence of a self-binding peptide | |||||||||
 マップデータ マップデータ | final map | |||||||||
 試料 試料 |
| |||||||||
 キーワード キーワード | inositol trisphosphate receptor / InsP3R / IP3R / cryoelectron microscopy / ion channel / calcium channel / isothermal titration calorimetry / self binding peptide / TRANSPORT PROTEIN | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報sensory perception of bitter taste / DAG and IP3 signaling / inositol 1,3,4,5 tetrakisphosphate binding / sensory perception of umami taste / inositol 1,4,5-trisphosphate-gated calcium channel activity / sensory perception of sweet taste / platelet dense tubular network membrane / Elevation of cytosolic Ca2+ levels / Effects of PIP2 hydrolysis / PLC beta mediated events ...sensory perception of bitter taste / DAG and IP3 signaling / inositol 1,3,4,5 tetrakisphosphate binding / sensory perception of umami taste / inositol 1,4,5-trisphosphate-gated calcium channel activity / sensory perception of sweet taste / platelet dense tubular network membrane / Elevation of cytosolic Ca2+ levels / Effects of PIP2 hydrolysis / PLC beta mediated events / inositol hexakisphosphate binding / inositol 1,4,5 trisphosphate binding / nuclear outer membrane / CLEC7A (Dectin-1) induces NFAT activation / intracellularly gated calcium channel activity / cytoplasmic side of endoplasmic reticulum membrane / brush border / Role of phospholipids in phagocytosis / calcium ion homeostasis / release of sequestered calcium ion into cytosol / Ion homeostasis / FCERI mediated Ca+2 mobilization / phosphatidylinositol binding / FCGR3A-mediated IL10 synthesis / secretory granule membrane / Antigen activates B Cell Receptor (BCR) leading to generation of second messengers / VEGFR2 mediated cell proliferation / sarcoplasmic reticulum / Regulation of insulin secretion / long-term synaptic potentiation / memory / Sensory perception of sweet, bitter, and umami (glutamate) taste / response to calcium ion / Glucagon-like Peptide-1 (GLP1) regulates insulin secretion / sensory perception of taste / apical part of cell / myelin sheath / Ca2+ pathway / positive regulation of cytosolic calcium ion concentration / receptor complex / G protein-coupled receptor signaling pathway / neuronal cell body / calcium ion binding / endoplasmic reticulum membrane / nucleolus / endoplasmic reticulum / nucleoplasm / membrane / plasma membrane / cytoplasm 類似検索 - 分子機能 | |||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | |||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.77 Å | |||||||||
 データ登録者 データ登録者 | Azumaya CM / Linton EA | |||||||||
| 資金援助 |  米国, 1件 米国, 1件
| |||||||||
 引用 引用 | ジャーナル: J Biol Chem / 年: 2020 タイトル: Cryo-EM structure of human type-3 inositol triphosphate receptor reveals the presence of a self-binding peptide that acts as an antagonist. 著者: Caleigh M Azumaya / Emily A Linton / Caitlin J Risener / Terunaga Nakagawa / Erkan Karakas / 要旨: Calcium-mediated signaling through inositol 1,4,5-triphosphate receptors (IPRs) is essential for the regulation of numerous physiological processes, including fertilization, muscle contraction, ...Calcium-mediated signaling through inositol 1,4,5-triphosphate receptors (IPRs) is essential for the regulation of numerous physiological processes, including fertilization, muscle contraction, apoptosis, secretion, and synaptic plasticity. Deregulation of IPRs leads to pathological calcium signaling and is implicated in many common diseases, including cancer and neurodegenerative, autoimmune, and metabolic diseases. Revealing the mechanism of activation and inhibition of this ion channel will be critical to an improved understanding of the biological processes that are controlled by IPRs. Here, we report structural findings of the human type-3 IPR (IPR-3) obtained by cryo-EM (at an overall resolution of 3.8 Å), revealing an unanticipated regulatory mechanism where a loop distantly located in the primary sequence occupies the IP-binding site and competitively inhibits IP binding. We propose that this inhibitory mechanism must differ qualitatively among IPR subtypes because of their diverse loop sequences, potentially serving as a key molecular determinant of subtype-specific calcium signaling in IPRs. In summary, our structural characterization of human IPR-3 provides critical insights into the mechanistic function of IPRs and into subtype-specific regulation of these important calcium-regulatory channels. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | EMマップ: SurfViewMolmilJmol/JSmol |
| 添付画像 |
- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_20849.map.gz | 139.2 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-20849-v30.xmlemd-20849.xml | 21.5 KB 21.5 KB | 表示 表示 | EMDBヘッダ |
| 画像 |  emd_20849.png emd_20849.png | 85.7 KB | ||
| マスクデータ | emd_20849_msk_1.map | 149.9 MB | マスクマップ | |
| Filedesc metadata | emd-20849.cif.gz | 7.6 KB | ||
| その他 | emd_20849_half_map_1.map.gzemd_20849_half_map_2.map.gz | 49 MB 49 MB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-20849ftp://ftp.pdbj.org/pub/emdb/structures/EMD-20849 http://ftp.pdbj.org/pub/emdb/structures/EMD-20849ftp://ftp.pdbj.org/pub/emdb/structures/EMD-20849 | HTTPS FTP |
-検証レポート
| 文書・要旨 | emd_20849_validation.pdf.gz | 990.1 KB | 表示 | EMDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | emd_20849_full_validation.pdf.gz | 989.7 KB | 表示 | |
| XML形式データ | emd_20849_validation.xml.gz | 14.7 KB | 表示 | |
| CIF形式データ | emd_20849_validation.cif.gz | 17.5 KB | 表示 | |
| アーカイブディレクトリ | https://ftp.pdbj.org/pub/emdb/validation_reports/EMD-20849ftp://ftp.pdbj.org/pub/emdb/validation_reports/EMD-20849 | HTTPS FTP |
-関連構造データ



-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| 「今月の分子」の関連する項目 |
















-マップ
| ファイル | ダウンロード / ファイル: emd_20849.map.gz / 形式: CCP4 / 大きさ: 149.9 MB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | final map | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ボクセルのサイズ | X=Y=Z: 1.247 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 密度 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 詳細 | EMDB XML:
CCP4マップ ヘッダ情報:
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
-添付データ
-マスク #1
| ファイル | emd_20849_msk_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
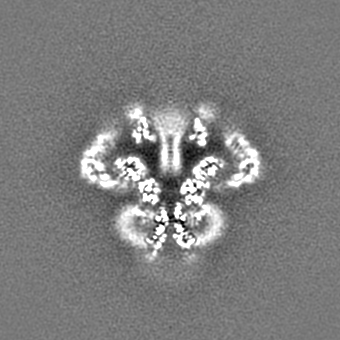
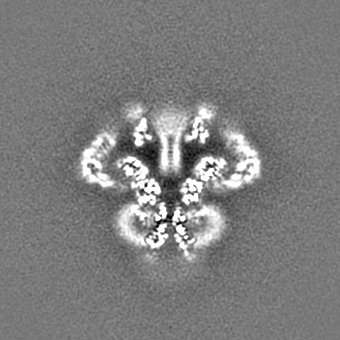
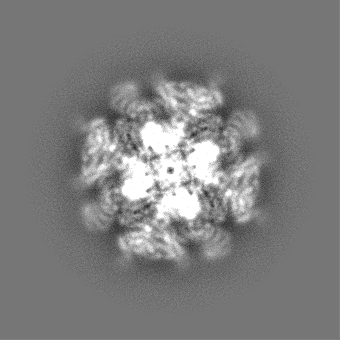
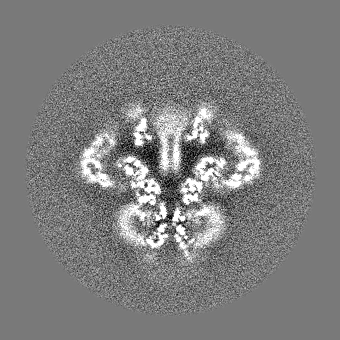
| 投影像・断面図 |
| ||||||||||||
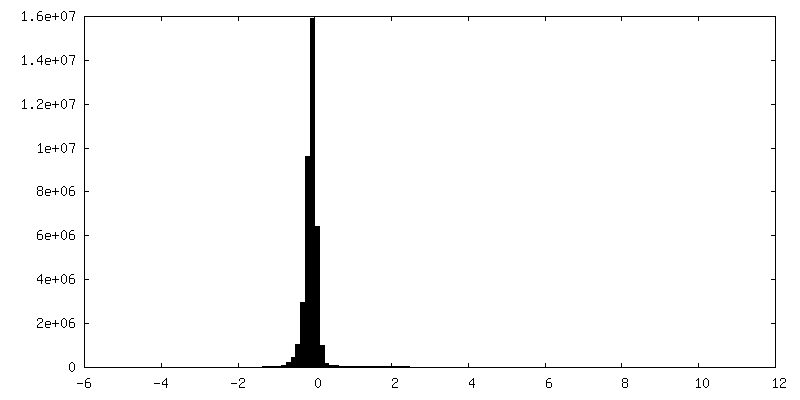
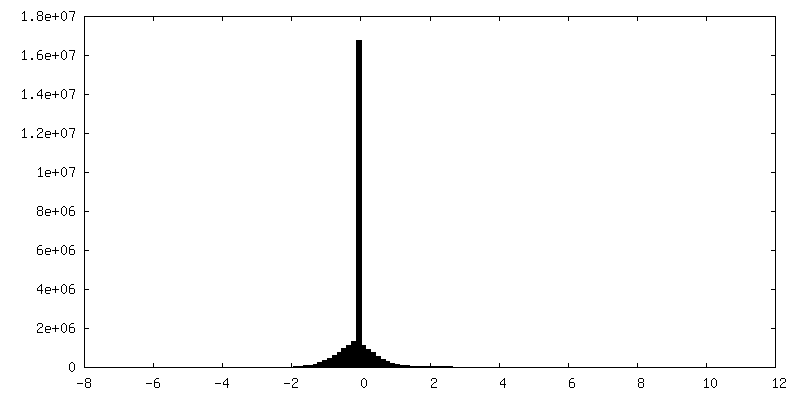
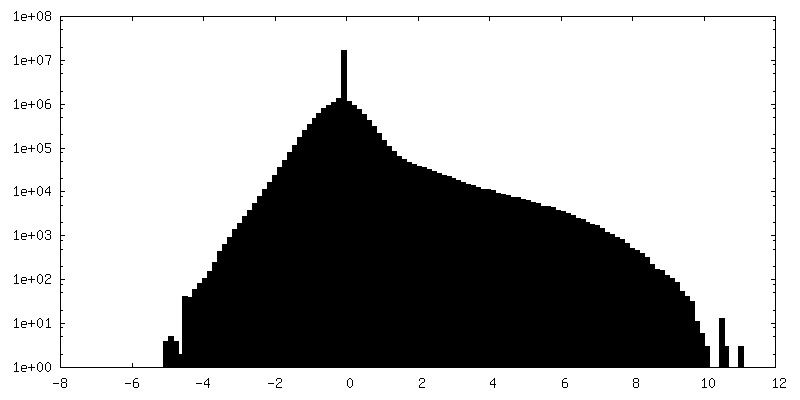
| 密度ヒストグラム |
 Z
Z Y
Y X
X




-ハーフマップ: #1
| ファイル | emd_20849_half_map_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
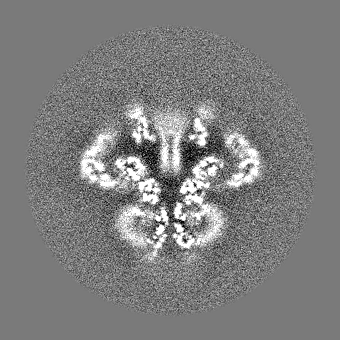
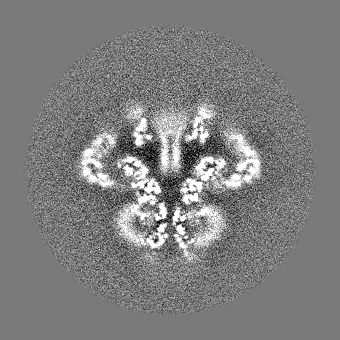
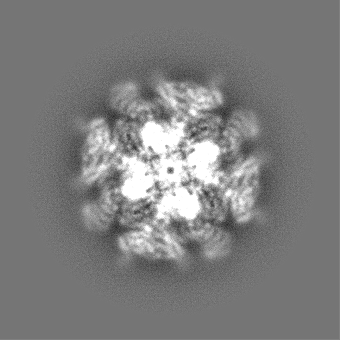
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |



-ハーフマップ: #2
| ファイル | emd_20849_half_map_2.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
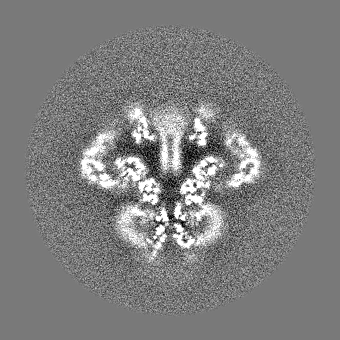
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |



- 試料の構成要素
試料の構成要素
-全体 : inositol 1,4,5-triphosphate receptor, type 3
| 全体 | 名称: inositol 1,4,5-triphosphate receptor, type 3 |
|---|---|
| 要素 |
|
-超分子 #1: inositol 1,4,5-triphosphate receptor, type 3
| 超分子 | 名称: inositol 1,4,5-triphosphate receptor, type 3 / タイプ: complex / ID: 1 / 親要素: 0 / 含まれる分子: #1 |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 1.2 MDa |
-分子 #1: inositol 1,4,5-triphosphate receptor, type 3
| 分子 | 名称: inositol 1,4,5-triphosphate receptor, type 3 / タイプ: protein_or_peptide / ID: 1 詳細: The full sequence of the sample is MSSFLHIGDIVSLYAEGSVNGFISTLGLVDDRCVVEPAAGDLDNPPKKFRDCLFKVCPMNRYSAQKQYWKAKQTKQDKEK ...詳細: The full sequence of the sample is MSSFLHIGDIVSLYAEGSVNGFISTLGLVDDRCVVEPAAGDLDNPPKKFRDCLFKVCPMNRYSAQKQYWKAKQTKQDKEK IADVVLLQKLQHAAQMEQKQNDTENKKVHGDVVKYGSVIQLLHMKSNKYLTVNKRLPALLEKNAMRVTLDATGNEGSWLF IQPFWKLRSNGDNVVVGDKVILNPVNAGQPLHASNYELSDNAGCKEVNSVNCNTSWKINLFMQFRDHLEEVLKGGDVVRL FHAEQEKFLTCDEYKGKLQVFLRTTLRQSATSATSSNALWEVEVVHHDPCRGGAGHWNGLYRFKHLATGNYLAAEENPSY KGDASDPKAAGMGAQGRTGRRNAGEKIKYCLVAVPHGNDIASLFELDPTTLQKTDSFVPRNSYVRLRHLCTNTWIQSTNV PIDIEEERPIRLMLGTCPTKEDKEAFAIVSVPVSEIRDLDFANDASSMLASAVEKLNEGFISQNDRRFVIQLLEDLVFFV SDVPNNGQNVLDIMVTKPNRERQKLMREQNILKQVFGILKAPFREKGGEGPLVRLEELSDQKNAPYQHMFRLCYRVLRHS QEDYRKNQEHIAKQFGMMQSQIGYDILAEDTITALLHNNRKLLEKHITKTEVETFVSLVRKNREPRFLDYLSDLCVSNHI AIPVTQELICKCVLDPKNSDILIRTELRPVKEMAQSHEYLSIEYSEEEVWLTWTDKNNEHHEKSVRQLAQEARAGNAHDE NVLSYYRYQLKLFARMCLDRQYLAIDEISQQLGVDLIFLCMADEMLPFDLRASFCHLMLHVHVDRDPQELVTPVKFARLW TEIPTAITIKDYDSNLNASRDDKKNKFANTMEFVEDYLNNVVSEAVPFANEEKNKLTFEVVSLAHNLIYFGFYSFSELLR LTRTLLGIIDCVQGPPAMLQAYEDPGGKNVRRSIQGVGHMMSTMVLSRKQSVFSAPSLSAGASAAEPLDRSKFEENEDIV VMETKLKILEILQFILNVRLDYRISYLLSVFKKEFVEVFPMQDSGADGTAPAFDSTTANMNLDRIGEQAEAMFGVGKTSS MLEVDDEGGRMFLRVLIHLTMHDYAPLVSGALQLLFKHFSQRQEAMHTFKQVQLLISAQDVENYKVIKSELDRLRTMVEK SELWVDKKGSGKGEEVEAGAAKDKKERPTDEEGFLHPPGEKSSENYQIVKGILERLNKMCGVGEQMRKKQQRLLKNMDAH KVMLDLLQIPYDKGDAKMMEILRYTHQFLQKFCAGNPGNQALLHKHLHLFLTPGLLEAETMQHIFLNNYQLCSEISEPVL QHFVHLLATHGRHVQYLDFLHTVIKAEGKYVKKCQDMIMTELTNAGDDVVVFYNDKASLAHLLDMMKAARDGVEDHSPLM YHISLVDLLAACAEGKNVYTEIKCTSLLPLEDVVSVVTHEDCITEVKMAYVNFVNHCYVDTEVEMKEIYTSNHIWTLFEN FTLDMARVCSKREKRVADPTLEKYVLSVVLDTINAFFSSPFSENSTSLQTHQTIVVQLLQSTTRLLECPWLQQQHKGSVE ACIRTLAMVAKGRAILLPMDLDAHISSMLSSGASCAAAAQRNASSYKATTRAFPRVTPTANQWDYKNIIEKLQDIITALE ERLKPLVQAELSVLVDVLHWPELLFLEGSEAYQRCESGGFLSKLIQHTKDLMESEEKLCIKVLRTLQQMLLKKTKYGDRG NQLRKMLLQNYLQNRKSTSRGDLPDPIGTGLDPDWSAIAATQCRLDKEGATKLVCDLITSTKNEKIFQESIGLAIHLLDG GNTEIQKSFHNLMMSDKKSERFFKVLHDRMKRAQQETKSTVAVNMNDLGSQPHEDREPVDPTTKGRVASFSIPGSSSRYS LGPSLRRGHEVSERVQSSEMGTSVLIMQPILRFLQLLCENHNRDLQNFLRCQNNKTNYNLVCETLQFLDIMCGSTTGGLG LLGLYINEDNVGLVIQTLETLTEYCQGPCHENQTCIVTHESNGIDIITALILNDISPLCKYRMDLVLQLKDNASKLLLAL MESRHDSENAERILISLRPQELVDVIKKAYLQEEERENSEVSPREVGHNIYILALQLSRHNKQLQHLLKPVKRIQEEEAE GISSMLSLNNKQLSQMLKSSAPAQEEEEDPLAYYENHTSQIEIVRQDRSMEQIVFPVPGICQFLTEETKHRLFTTTEQDE QGSKVSDFFDQSSFLHNEMEWQRKLRSMPLIYWFSRRMTLWGSISFNLAVFINIIIAFFYPYMEGASTGVLDSPLISLLF WILICFSIAALFTKRYSIRPLIVALILRSIYYLGIGPTLNILGALNLTNKIVFVVSFVGNRGTFIRGYKAMVMDMEFLYH VGYILTSVLGLFAHELFYSILLFDLIYREETLFNVIKSVTRNGRSILLTALLALILVYLFSIVGFLFLKDDFILEVDRLP NNHSTASPLGMPHGAAAFVDTCSGDKMDCVSGLSVPEVLEEDRELDSTERACDTLLMCIVTVMNHGLRNGGGVGDILRKP SKDESLFPARVVYDLLFFFIVIIIVLNLIFGVIIDTFADLRSEKQKKEEILKTTCFICGLERDKFDNKTVSFEEHIKLEH NMWNYLYFIVLVRVKNKTDYTGPESYVAQMIKNKNLDWFPRMRAMSLVSNEGEGEQNEIRILQDKLNSTMKLVSHLTAQL NELKEQMTEQRKRRQRLGFVDVQNCISRGENLYFQSAWSHPQFEKGGGSGGGSGGSAWSHPQFEK コピー数: 4 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 278.584406 KDa |
| 組換発現 | 生物種:   Spodoptera frugiperda (ツマジロクサヨトウ) Spodoptera frugiperda (ツマジロクサヨトウ) |
| 配列 | 文字列: MSSFLHIGDI VSLYAEGSVN GFISTLGLVD DRCVVEPAAG DLDNPPKKFR DCLFKVCPMN RYSAQKQYWK AKQTKQDKEK IADVVLLQK LQHAAQMEQK QNDTENKKVH GDVVKYGSVI QLLHMKSNKY LTVNKRLPAL LEKNAMRVTL DATGNEGSWL F IQPFWKLR ...文字列: MSSFLHIGDI VSLYAEGSVN GFISTLGLVD DRCVVEPAAG DLDNPPKKFR DCLFKVCPMN RYSAQKQYWK AKQTKQDKEK IADVVLLQK LQHAAQMEQK QNDTENKKVH GDVVKYGSVI QLLHMKSNKY LTVNKRLPAL LEKNAMRVTL DATGNEGSWL F IQPFWKLR SNGDNVVVGD KVILNPVNAG QPLHASNYEL SDNAGCKEVN SVNCNTSWKI NLFMQFRDHL EEVLKGGDVV RL FHAEQEK FLTCDEYKGK LQVFLRTTLR QSATSATSSN ALWEVEVVHH DPCRGGAGHW NGLYRFKHLA TGNYLAAEEN PSY KGDASD PKAAGMGAQG RTGRRNAGEK IKYCLVAVPH GNDIASLFEL DPTTLQKTDS FVPRNSYVRL RHLCTNTWIQ STNV PIDIE EERPIRLMLG TCPTKEDKEA FAIVSVPVSE IRDLDFANDA SSMLASAVEK LNEGFISQND RRFVIQLLED LVFFV SDVP NNGQNVLDIM VTKPNRERQK LMREQNILKQ VFGILKAPFR EKGGEGPLVR LEELSDQKNA PYQHMFRLCY RVLRHS QED YRKNQEHIAK QFGMMQSQIG YDILAEDTIT ALLHNNRKLL EKHITKTEVE TFVSLVRKNR EPRFLDYLSD LCVSNHI AI PVTQELICKC VLDPKNSDIL IRTELRPVKE MAQSHEYLSI EYSEEEVWLT WTDKNNEHHE KSVRQLAQEA RAGNAHDE N VLSYYRYQLK LFARMCLDRQ YLAIDEISQQ LGVDLIFLCM ADEMLPFDLR ASFCHLMLHV HVDRDPQELV TPVKFARLW TEIPTAITIK DYDSNLNASR DDKKNKFANT MEFVEDYLNN VVSEAVPFAN EEKNKLTFEV VSLAHNLIYF GFYSFSELLR LTRTLLGII DCVQGPPAML QAYEDPGGKN VRRSIQGVGH MMSTMVLSRK QSVFSAPSLS AGASAAEPLD RSKFEENEDI V VMETKLKI LEILQFILNV RLDYRISYLL SVFKKEFVEV FPMQDSGADG TAPAFDSTTA NMNLDRIGEQ AEAMFGVGKT SS MLEVDDE GGRMFLRVLI HLTMHDYAPL VSGALQLLFK HFSQRQEAMH TFKQVQLLIS AQDVENYKVI KSELDRLRTM VEK SELWVD KKGSGKGEEV EAGAAKDKKE RPTDEEGFLH PPGEKSSENY QIVKGILERL NKMCGVGEQM RKKQQRLLKN MDAH KVMLD LLQIPYDKGD AKMMEILRYT HQFLQKFCAG NPGNQALLHK HLHLFLTPGL LEAETMQHIF LNNYQLCSEI SEPVL QHFV HLLATHGRHV QYLDFLHTVI KAEGKYVKKC QDMIMTELTN AGDDVVVFYN DKASLAHLLD MMKAARDGVE DHSPLM YHI SLVDLLAACA EGKNVYTEIK CTSLLPLEDV VSVVTHEDCI TEVKMAYVNF VNHC(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)DYKNIIEKL QDIITALEER L KPLVQAEL SVLVDVLHWP ELLFLEGSEA YQRCESGGFL SKLIQHTKDL MESEEKLCIK VLRTLQQMLL KKTKYGDRGN QL RKMLLQN YLQNRKSTSR GDLPDPIGTG LDPDWSAIAA TQCRLDKEGA TKLVCDLITS TKNEKIFQES IGLAIHLLDG GNT EIQKSF HNLMMSDKKS ERFFKVLHDR MKRAQQETKS TVAVNMNDLG SQPHEDREPV DPTTKGRVAS FSIPGSSSRY SLGP SLRRG HEVSERVQSS EMGTSVLIMQ PILRFLQLLC ENHNRDLQNF LRCQNNKTNY NLVCETLQFL DIMCGSTTGG LGLLG LYIN EDNVGLVIQT LETLTEYCQG PCHENQTCIV THESNGIDII TALILNDISP LCKYRMDLVL QLKDNASKLL LALMES RHD SENAERILIS LRPQELVDVI KKAYLQEEER ENSEVSPREV GHNIYILALQ LSRHNKQLQH LLKPVKRIQE EEAEGIS SM LSLNNKQLSQ MLKSSAPAQE EEEDPLAYYE NHTSQIEIVR QDRSMEQIVF PVPGICQFLT EETKHRLFTT TEQDEQGS K VSDFFDQSSF LHNEMEWQRK LRSMPLIYWF SRRMTLWGSI SFNLAVFINI IIAFFYPYME (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) PLIVALILRS IYYLGIGPTL NILGALNLTN KIVFVVSFVG NRGTFIRGYK AMVMDMEFL YHVGYILTSV LGLFAHELFY SILLFDLIYR EETLFNVIKS VTRNGRSILL TALLALILVY LFSIVGFLFL K DDFILEVD RLPNNHSTAS PLGMPHGAAA FVDTCSGDKM DCVSGLSVPE VLEEDRELDS TERACDTLLM CIVTVMNHGL RN GGGVGDI LRKPSKDESL FPARVVYDLL FFFIVIIIVL NLIFGVIIDT FADLRSEKQK KEEILKTTCF ICGLERDKFD NKT VSFEEH IKLEHNMWNY LYFIVLVRVK NKTDYTGPES YVAQMIKNKN LDWFPRMRAM SLV(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK) |
-分子 #2: ZINC ION
| 分子 | 名称: ZINC ION / タイプ: ligand / ID: 2 / コピー数: 4 / 式: ZN |
|---|---|
| 分子量 | 理論値: 65.409 Da |
-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | particle |
-試料調製
| 濃度 | 1.3 mg/mL | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 緩衝液 | pH: 8 構成要素:
| ||||||||||||||
| グリッド | モデル: C-flat-2/1 / 材質: COPPER / メッシュ: 200 / 前処理 - タイプ: GLOW DISCHARGE / 前処理 - 時間: 120 sec. / 詳細: 25 mA | ||||||||||||||
| 凍結 | 凍結剤: ETHANE / チャンバー内湿度: 100 % / チャンバー内温度: 281 K / 装置: FEI VITROBOT MARK IV / 詳細: The grid was blotted for 3 seconds at force 1. |
- 電子顕微鏡法
電子顕微鏡法
| 顕微鏡 | FEI POLARA 300 |
|---|---|
| 撮影 | フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 検出モード: COUNTING / 平均電子線量: 70.0 e/Å2 |
| 電子線 | 加速電圧: 300 kV / 電子線源:  FIELD EMISSION GUN FIELD EMISSION GUN |
| 電子光学系 | 照射モード: FLOOD BEAM / 撮影モード: BRIGHT FIELD / Cs: 2.2 mm / 倍率(公称値): 31000 |
| 試料ステージ | ホルダー冷却材: NITROGEN |
| 実験機器 |  モデル: Tecnai Polara / 画像提供: FEI Company |
-画像解析
| 初期モデル | モデルのタイプ: EMDB MAP EMDB ID: |
|---|---|
| 最終 再構成 | 想定した対称性 - 点群: C4 (4回回転対称) / 解像度のタイプ: BY AUTHOR / 解像度: 3.77 Å / 解像度の算出法: FSC 0.143 CUT-OFF / ソフトウェア - 名称: cisTEM / 使用した粒子像数: 82511 |
| 初期 角度割当 | タイプ: MAXIMUM LIKELIHOOD |
| 最終 角度割当 | タイプ: MAXIMUM LIKELIHOOD |
