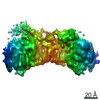
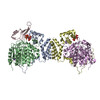
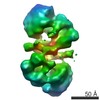
登録情報 データベース : PDB / ID : 7ao8タイトル Structure of the MTA1/HDAC1/MBD2 NURD deacetylase complex Histone deacetylase 1 Metastasis-associated protein MTA1 Methyl-CpG-binding domain protein 2 キーワード / / 機能・相同性 分子機能 ドメイン・相同性 構成要素
/ / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / 生物種 Homo sapiens (ヒト)手法 / / / 解像度 : 4.5 Å データ登録者 Millard, C.J. / Fairall, L. / Ragan, T.J. / Savva, C.G. / Schwabe, J.W.R. 資金援助 組織 認可番号 国 Wellcome Trust WT100237/Z/12/Z Medical Research Council (MRC, United Kingdom) MC_PC_17136 Wolfson Foundation
ジャーナル : Nucleic Acids Res / 年 : 2020タイトル : The topology of chromatin-binding domains in the NuRD deacetylase complex.著者 : Christopher J Millard / Louise Fairall / Timothy J Ragan / Christos G Savva / John W R Schwabe / 要旨 : Class I histone deacetylase complexes play essential roles in many nuclear processes. Whilst they contain a common catalytic subunit, they have diverse modes of action determined by associated ... Class I histone deacetylase complexes play essential roles in many nuclear processes. Whilst they contain a common catalytic subunit, they have diverse modes of action determined by associated factors in the distinct complexes. The deacetylase module from the NuRD complex contains three protein domains that control the recruitment of chromatin to the deacetylase enzyme, HDAC1/2. Using biochemical approaches and cryo-electron microscopy, we have determined how three chromatin-binding domains (MTA1-BAH, MBD2/3 and RBBP4/7) are assembled in relation to the core complex so as to facilitate interaction of the complex with the genome. We observe a striking arrangement of the BAH domains suggesting a potential mechanism for binding to di-nucleosomes. We also find that the WD40 domains from RBBP4 are linked to the core with surprising flexibility that is likely important for chromatin engagement. A single MBD2 protein binds asymmetrically to the dimerisation interface of the complex. This symmetry mismatch explains the stoichiometry of the complex. Finally, our structures suggest how the holo-NuRD might assemble on a di-nucleosome substrate. 履歴 登録 2020年10月14日 登録サイト / 処理サイト 改定 1.0 2020年11月11日 Provider / タイプ 改定 1.1 2020年12月30日 Group / カテゴリ / citation_authorItem _citation.country / _citation.journal_abbrev ... _citation.country / _citation.journal_abbrev / _citation.journal_id_ASTM / _citation.journal_id_CSD / _citation.journal_id_ISSN / _citation.journal_volume / _citation.page_first / _citation.page_last / _citation.pdbx_database_id_DOI / _citation.pdbx_database_id_PubMed / _citation.title / _citation.year / _citation_author.identifier_ORCID 改定 1.2 2024年5月1日 Group / Database references / Refinement descriptionカテゴリ chem_comp_atom / chem_comp_bond ... chem_comp_atom / chem_comp_bond / database_2 / em_3d_fitting_list / pdbx_initial_refinement_model Item _database_2.pdbx_DOI / _database_2.pdbx_database_accession ... _database_2.pdbx_DOI / _database_2.pdbx_database_accession / _em_3d_fitting_list.accession_code / _em_3d_fitting_list.initial_refinement_model_id / _em_3d_fitting_list.source_name / _em_3d_fitting_list.type
すべて表示 表示を減らす
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード TRANSCRIPTION (転写 (生物学)) /
TRANSCRIPTION (転写 (生物学)) /  機能・相同性情報
機能・相同性情報
 データ登録者
データ登録者 英国, 3件
英国, 3件  引用
引用 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード







 PDBj
PDBj































 集合体
集合体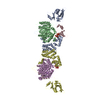
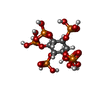


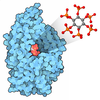
 分子量: 660.035 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C6H18O24P6
分子量: 660.035 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C6H18O24P6 分子量: 65.409 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Zn
分子量: 65.409 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Zn 分子量: 39.098 Da / 分子数: 4 / 由来タイプ: 合成 / 式: K
分子量: 39.098 Da / 分子数: 4 / 由来タイプ: 合成 / 式: K 試料調製
試料調製 電子顕微鏡撮影
電子顕微鏡撮影
 解析
解析

