 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 5jhn | ||||||
|---|---|---|---|---|---|---|---|
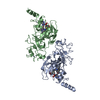
| タイトル | Structure of G9a SET-domain with Histone H3K9Ala mutant peptide and bound S-adenosylmethionine | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード |  TRANSFERASE (転移酵素) / SET-domain / Histone methyl transferase (ヒストンメチルトランスフェラーゼ) / histone peptide TRANSFERASE (転移酵素) / SET-domain / Histone methyl transferase (ヒストンメチルトランスフェラーゼ) / histone peptide | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報regulation of protein modification process / histone H3K56 methyltransferase activity / : / phenotypic switching / neuron fate specification / : / [histone H3]-lysine9 N-methyltransferase / histone H3K27 methyltransferase activity / histone H3K9 methyltransferase activity / histone H3K9me2 methyltransferase activity ...regulation of protein modification process / histone H3K56 methyltransferase activity / : / phenotypic switching / neuron fate specification / : / [histone H3]-lysine9 N-methyltransferase / histone H3K27 methyltransferase activity / histone H3K9 methyltransferase activity / histone H3K9me2 methyltransferase activity / peptidyl-lysine dimethylation / synaptonemal complex assembly / negative regulation of autophagosome assembly / oocyte development / protein-lysine N-methyltransferase activity / C2H2 zinc finger domain binding / 受精 / : / cellular response to cocaine / : / organ growth / Transcriptional Regulation by E2F6 / spermatid development / behavioral response to cocaine / regulation of DNA replication / RNA Polymerase I Transcription Initiation / Transcriptional Regulation by VENTX / long-term memory / Chromatin modifying enzymes / epigenetic regulation of gene expression / response to fungicide / 転移酵素; 一炭素原子の基を移すもの; メチル基を移すもの / telomere organization / cellular response to starvation / RNA Polymerase I Promoter Opening / Interleukin-7 signaling / Assembly of the ORC complex at the origin of replication / DNAメチル化 / Condensation of Prophase Chromosomes / transcription corepressor binding / HCMV Late Events / Chromatin modifications during the maternal to zygotic transition (MZT) / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / SIRT1 negatively regulates rRNA expression / PRC2 methylates histones and DNA / Defective pyroptosis / promoter-specific chromatin binding / HDACs deacetylate histones / RNA Polymerase I Promoter Escape / RNA polymerase II transcription regulatory region sequence-specific DNA binding / Transcriptional regulation by small RNAs / Formation of the beta-catenin:TCF transactivating complex / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / NoRC negatively regulates rRNA expression / B-WICH complex positively regulates rRNA expression / HDMs demethylate histones / Regulation of TP53 Activity through Methylation / PKMTs methylate histone lysines / RMTs methylate histone arginines / 遺伝的組換え / Pre-NOTCH Transcription and Translation / nucleosome assembly / Activation of anterior HOX genes in hindbrain development during early embryogenesis / HCMV Early Events / Transcriptional regulation of granulopoiesis / structural constituent of chromatin / ヌクレオソーム / cellular response to xenobiotic stimulus / p53 binding / 遺伝子発現 / RUNX1 regulates transcription of genes involved in differentiation of HSCs / chromatin organization / Factors involved in megakaryocyte development and platelet production / HATs acetylate histones / Senescence-Associated Secretory Phenotype (SASP) / response to ethanol / Oxidative Stress Induced Senescence / Estrogen-dependent gene expression / nuclear speck / cadherin binding / Amyloid fiber formation / protein heterodimerization activity / クロマチン / negative regulation of transcription by RNA polymerase II / protein-containing complex / DNA binding / extracellular exosome / zinc ion binding / extracellular region / 核質 / 生体膜 / 細胞核類似検索 - 分子機能 | ||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||
| 手法 | X線回折 / シンクロトロン / 多波長異常分散 / 解像度: 1.67 Å | ||||||
 データ登録者 データ登録者 | Jayaram, H. / Bellon, S.F. / Poy, F. | ||||||
 引用 引用 | ジャーナル: Proc.Natl.Acad.Sci.USA / 年: 2016 タイトル: S-adenosyl methionine is necessary for inhibition of the methyltransferase G9a by the lysine 9 to methionine mutation on histone H3. 著者: Jayaram, H. / Hoelper, D. / Jain, S.U. / Cantone, N. / Lundgren, S.M. / Poy, F. / Allis, C.D. / Cummings, R. / Bellon, S. / Lewis, P.W. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 5jhn.cif.gz | 132.1 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb5jhn.ent.gz | 101.8 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 5jhn.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/jh/5jhnftp://data.pdbj.org/pub/pdb/validation_reports/jh/5jhn | HTTPS FTP |
|---|
-関連構造データ
-リンク
 PDBj
PDBj











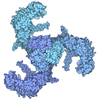





- 集合体
集合体
| 登録構造単位 | 
| ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||||||||||||
| 単位格子 |
| ||||||||||||||||||
| 非結晶学的対称性 (NCS) | NCSドメイン:
NCSドメイン領域: Component-ID: 0 / Ens-ID: 1 / Beg auth comp-ID: GLU / Beg label comp-ID: GLU / End auth comp-ID: ARG / End label comp-ID: ARG / Refine code: 0 / Auth seq-ID: 919 - 1188 / Label seq-ID: 4 - 273
|
-要素
| #1: タンパク質 | 分子量: 31661.904 Da / 分子数: 2 / 断片: UNP residues 882-1155 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: EHMT2, BAT8, C6orf30, G9A, KMT1C, NG36 / 発現宿主:  Escherichia coli (大腸菌) / 株 (発現宿主): BL21(DE3) Rosetta Escherichia coli (大腸菌) / 株 (発現宿主): BL21(DE3) Rosetta参照: UniProt: Q96KQ7, 転移酵素; 一炭素原子の基を移すもの; メチル基を移すもの, ヒストンメチルトランスフェラーゼ#2: タンパク質・ペプチド | 分子量: 1079.166 Da / 分子数: 2 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) / 参照: UniProt: P68431*PLUS#3: 化合物 | ChemComp-ZN /   分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn#4: 化合物 | S-アデノシルメチオニン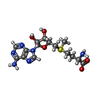  分子量: 398.437 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C15H22N6O5S 分子量: 398.437 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C15H22N6O5S#5: 水 | ChemComp-HOH / | 水 分子量: 18.015 Da / 分子数: 154 / 由来タイプ: 天然 / 式: H2O 分子量: 18.015 Da / 分子数: 154 / 由来タイプ: 天然 / 式: H2O |
|---|
-実験情報
-実験
| 実験 | 手法: X線回折 / 使用した結晶の数: 1 |
|---|
- 試料調製
試料調製
| 結晶 | マシュー密度: 2.49 Å3/Da / 溶媒含有率: 50.52 % |
|---|---|
| 結晶化 | 温度: 298 K / 手法: 蒸気拡散法 / pH: 7 詳細: 0.2 M Tri-ammonium citrate pH 7 and 20% w/v polyethylene glycol 3350 |
-データ収集
| 回折 | 平均測定温度: 175 K |
|---|---|
| 放射光源 | 由来: シンクロトロン / サイト: APS  / ビームライン: 14-BM-C / 波長: 1.01 Å / ビームライン: 14-BM-C / 波長: 1.01 Å |
| 検出器 | タイプ: ADSC QUANTUM 210 / 検出器: CCD / 日付: 2012年1月1日 |
| 放射 | プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray |
| 放射波長 | 波長: 1.01 Å / 相対比: 1 |
| 反射 | 解像度: 1.67→34.5 Å / Num. obs: 70175 / % possible obs: 99.19 % / 冗長度: 3.7 % / Net I/σ(I): 16.67 |
- 解析
解析
| ソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 精密化 | 構造決定の手法: 多波長異常分散 / 解像度: 1.67→34.5 Å / Cor.coef. Fo:Fc: 0.963 / Cor.coef. Fo:Fc free: 0.952 / SU B: 2.882 / SU ML: 0.092 / 交差検証法: THROUGHOUT / ESU R: 0.108 / ESU R Free: 0.105 / 詳細: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | イオンプローブ半径: 0.8 Å / 減衰半径: 0.8 Å / VDWプローブ半径: 1.2 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 36.414 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: 1 / 解像度: 1.67→34.5 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
|