ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 9okc | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
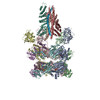
| タイトル | 22bin20S complex (NSF-alphaSNAP-2:2 syntaxin-1a:SNAP-25), hydrolyzing, class 17 | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | HYDROLASE / ATPase / SNARE / hydrolysis / disassembly / translocation / exocytosis / neurotransmitter release / synapse / synaptic transmission / membrane fusion | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報SNARE complex disassembly / ATP-dependent protein disaggregase activity / intra-Golgi vesicle-mediated transport / Golgi to plasma membrane protein transport / Golgi stack / vesicle-fusing ATPase / syntaxin-1 binding / positive regulation of receptor recycling / ionotropic glutamate receptor binding / SNARE binding ...SNARE complex disassembly / ATP-dependent protein disaggregase activity / intra-Golgi vesicle-mediated transport / Golgi to plasma membrane protein transport / Golgi stack / vesicle-fusing ATPase / syntaxin-1 binding / positive regulation of receptor recycling / ionotropic glutamate receptor binding / SNARE binding / PDZ domain binding / intracellular protein transport / potassium ion transport / positive regulation of protein catabolic process / midbody / protein kinase binding / protein-containing complex binding / ATP hydrolysis activity / ATP binding / metal ion binding / identical protein binding / plasma membrane / cytosol 類似検索 - 分子機能 | ||||||||||||
| 生物種 |   Cricetulus griseus (モンゴルキヌゲネズミ) Cricetulus griseus (モンゴルキヌゲネズミ) | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.67 Å | ||||||||||||
 データ登録者 データ登録者 | White, K.I. / Brunger, A.T. | ||||||||||||
| 資金援助 |  米国, 3件 米国, 3件
| ||||||||||||
 引用 引用 | ジャーナル: Nat Commun / 年: 2025 タイトル: Structural remodeling of target-SNARE protein complexes by NSF enables synaptic transmission. 著者: K Ian White / Yousuf A Khan / Kangqiang Qiu / Ashwin Balaji / Sergio Couoh-Cardel / Luis Esquivies / Richard A Pfuetzner / Jiajie Diao / Axel T Brunger / 要旨: Synaptic vesicles containing neurotransmitters fuse with the plasma membrane upon the arrival of an action potential at the active zone. Multiple proteins organize trans-SNARE complex assembly and ...Synaptic vesicles containing neurotransmitters fuse with the plasma membrane upon the arrival of an action potential at the active zone. Multiple proteins organize trans-SNARE complex assembly and priming, leading to fusion. One target membrane SNARE, syntaxin, forms nanodomains at the active zone, and another, SNAP-25, enters non-fusogenic complexes with it. Here, we reveal mechanistic details of AAA+ protein NSF (N-ethylmaleimide sensitive factor) and SNAP (soluble NSF attachment protein) action before fusion. We show that syntaxin clusters are conserved, that NSF colocalizes with them, and characterize SNARE populations that may exist within or near them using cryo-EM. Supercomplexes of NSF, α-SNAP, and either a syntaxin tetramer or one of two binary complexes of syntaxin-SNAP-25 reveal atomic details of SNARE processing and show how sequential ATP hydrolysis drives disassembly. These results suggest a functional role for syntaxin clusters as reservoirs and a corresponding role for NSF in syntaxin liberation and SNARE protein quality control preceding fusion. #1: ジャーナル: bioRxiv / 年: 2025 タイトル: Structural remodeling of target-SNARE protein complexes by NSF enables synaptic transmission. 著者: K Ian White / Yousuf A Khan / Kangqiang Qiu / Ashwin Balaji / Sergio Couoh-Cardel / Luis Esquivies / Richard A Pfuetzner / Jiajie Diao / Axel T Brunger / 要旨: Synaptic vesicles containing neurotransmitters fuse with the plasma membrane upon the arrival of an action potential at the active zone. Multiple proteins organize trans-SNARE complex assembly and ...Synaptic vesicles containing neurotransmitters fuse with the plasma membrane upon the arrival of an action potential at the active zone. Multiple proteins organize trans-SNARE complex assembly and priming, leading to fusion. One target membrane SNARE, syntaxin, forms nanodomains at the active zone, and another, SNAP-25, enters non-fusogenic complexes with it. Here, we reveal mechanistic details of AAA+ protein NSF (N-ethylmaleimide sensitive factor) and SNAP (soluble NSF attachment protein) action before fusion. We show that syntaxin clusters are conserved, that NSF colocalizes with them, and characterize SNARE populations that may exist within or near them using cryo-EM. Supercomplexes of NSF, α-SNAP, and either a syntaxin tetramer or one of two binary complexes of syntaxin-SNAP-25 reveal atomic details of SNARE processing and show how sequential ATP hydrolysis drives disassembly. These results suggest a functional role for syntaxin clusters as reservoirs and a corresponding role for NSF in syntaxin liberation and SNARE protein quality control preceding fusion. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 9okc.cif.gz | 1 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb9okc.ent.gz | 875.8 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 9okc.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 9okc_validation.pdf.gz | 2.3 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 9okc_full_validation.pdf.gz | 2.3 MB | 表示 | |
| XML形式データ | 9okc_validation.xml.gz | 92 KB | 表示 | |
| CIF形式データ | 9okc_validation.cif.gz | 141.1 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/ok/9okcftp://data.pdbj.org/pub/pdb/validation_reports/ok/9okc | HTTPS FTP |
-関連構造データ
| 関連構造データ |  70559MC  9ojrC  9ojuC  9ojzC  9ok3C  9ok5C  9oljC  9oloC  9om6C  9omqC  9pafC  9pagC  9pb9C  9pbaC  9pbfC  9pbvC  9pc3C  9pcxC  9pczC  9pd1C  9pd8C  9pdbC  9pddC  9pf2C  9pfcC  9pffC  9pfgC  9oj2 9ojj C: 同じ文献を引用 ( M: このデータのモデリングに利用したマップデータ |
|---|---|
| 類似構造データ |


























-リンク
 PDBj
PDBj


- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
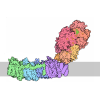
-タンパク質 / タンパク質・ペプチド , 2種, 7分子 ABCDEFG
| #1: タンパク質 | 分子量: 82907.430 Da / 分子数: 6 / 由来タイプ: 組換発現 由来: (組換発現) Cricetulus griseus (モンゴルキヌゲネズミ)遺伝子: NSF / 発現宿主:  #2: タンパク質・ペプチド | | 分子量: 1039.273 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) |
|---|
-非ポリマー , 4種, 17分子 

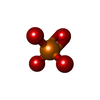

| #3: 化合物 | ChemComp-ADP /  分子量: 427.201 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / タイプ: SUBJECT OF INVESTIGATION / コメント: ADP, エネルギー貯蔵分子*YM 分子量: 427.201 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / タイプ: SUBJECT OF INVESTIGATION / コメント: ADP, エネルギー貯蔵分子*YM#4: 化合物 | ChemComp-ATP /  分子量: 507.181 Da / 分子数: 8 / 由来タイプ: 合成 / 式: C10H16N5O13P3 / タイプ: SUBJECT OF INVESTIGATION / コメント: ATP, エネルギー貯蔵分子*YM 分子量: 507.181 Da / 分子数: 8 / 由来タイプ: 合成 / 式: C10H16N5O13P3 / タイプ: SUBJECT OF INVESTIGATION / コメント: ATP, エネルギー貯蔵分子*YM#5: 化合物 |  分子量: 94.971 Da / 分子数: 3 / 由来タイプ: 合成 / 式: PO4 / タイプ: SUBJECT OF INVESTIGATION 分子量: 94.971 Da / 分子数: 3 / 由来タイプ: 合成 / 式: PO4 / タイプ: SUBJECT OF INVESTIGATION#6: 化合物 |  分子量: 24.305 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION 分子量: 24.305 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | Y |
|---|---|
| Has protein modification | N |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 実験値: NO | ||||||||||||||||||||||||||||||
| 由来(天然) |
| ||||||||||||||||||||||||||||||
| 由来(組換発現) |
| ||||||||||||||||||||||||||||||
| 緩衝液 | pH: 8 | ||||||||||||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||||||||||||
| 試料 | 濃度: 15 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||
| 試料支持 | 詳細: 15 mA in PELCO easiGlow Glow Discharge Cleaning System グリッドの材料: GOLD / グリッドのサイズ: 200 divisions/in. / グリッドのタイプ: Quantifoil R1.2/1.3 | ||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 298 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: TFS KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 22500 X / 最大 デフォーカス(公称値): 3000 nm / 最小 デフォーカス(公称値): 1000 nm / Cs: 2.7 mm / アライメント法: COMA FREE |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 電子線照射量: 33.96 e/Å2 / フィルム・検出器のモデル: GATAN K3 (6k x 4k) |
- 解析
解析
| EMソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 1889717 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.67 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 21214 / アルゴリズム: BACK PROJECTION / 対称性のタイプ: POINT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: FLEXIBLE FIT / 空間: REAL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | PDB-ID: 6MDM Accession code: 6MDM / Source name: PDB / タイプ: experimental model |