 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 9j0o | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
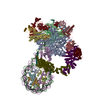
| タイトル | Arrested elongation complex of mammalian RNA polymerase II with nucleosome (AEC1-nuc) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 要素 要素 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 キーワード キーワード | TRANSCRIPTION / RNA polymerase / NELF / nucleosome / DNA / RNA | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報NELF complex / NTRK3 as a dependence receptor / negative regulation of DNA-templated transcription, elongation / negative regulation of chromosome condensation / Barr body / DSIF complex / : / regulation of transcription elongation by RNA polymerase II / inner kinetochore / pericentric heterochromatin formation ...NELF complex / NTRK3 as a dependence receptor / negative regulation of DNA-templated transcription, elongation / negative regulation of chromosome condensation / Barr body / DSIF complex / : / regulation of transcription elongation by RNA polymerase II / inner kinetochore / pericentric heterochromatin formation / Formation of RNA Pol II elongation complex / Formation of the Early Elongation Complex / Transcriptional regulation by small RNAs / RNA Polymerase II Pre-transcription Events / TP53 Regulates Transcription of DNA Repair Genes / FGFR2 alternative splicing / RNA polymerase II transcribes snRNA genes / mRNA Capping / mRNA Splicing - Minor Pathway / Processing of Capped Intron-Containing Pre-mRNA / RNA Polymerase II Promoter Escape / RNA Polymerase II Transcription Pre-Initiation And Promoter Opening / RNA Polymerase II Transcription Initiation / RNA Polymerase II Transcription Elongation / RNA Polymerase II Transcription Initiation And Promoter Clearance / RNA Pol II CTD phosphorylation and interaction with CE / Estrogen-dependent gene expression / Formation of TC-NER Pre-Incision Complex / Dual incision in TC-NER / Gap-filling DNA repair synthesis and ligation in TC-NER / mRNA Splicing - Major Pathway / negative regulation of stem cell differentiation / muscle cell differentiation / nuclear lumen / positive regulation of DNA-templated transcription, elongation / oocyte maturation / Abortive elongation of HIV-1 transcript in the absence of Tat / nucleosomal DNA binding / transcription elongation-coupled chromatin remodeling / RNA Pol II CTD phosphorylation and interaction with CE during HIV infection / RNA Pol II CTD phosphorylation and interaction with CE / nucleus organization / Formation of the Early Elongation Complex / Formation of the HIV-1 Early Elongation Complex / mRNA Capping / RNA polymerase II complex binding / maintenance of transcriptional fidelity during transcription elongation by RNA polymerase II / Pausing and recovery of Tat-mediated HIV elongation / Tat-mediated HIV elongation arrest and recovery / positive regulation of macroautophagy / negative regulation of transcription elongation by RNA polymerase II / RNA polymerase II transcribes snRNA genes / HIV elongation arrest and recovery / Pausing and recovery of HIV elongation / spermatid development / negative regulation of tumor necrosis factor-mediated signaling pathway / Tat-mediated elongation of the HIV-1 transcript / single fertilization / Formation of HIV-1 elongation complex containing HIV-1 Tat / subtelomeric heterochromatin formation / RNA polymerase I complex / RNA polymerase III complex / Formation of HIV elongation complex in the absence of HIV Tat / negative regulation of megakaryocyte differentiation / RNA polymerase II core promoter sequence-specific DNA binding / protein localization to CENP-A containing chromatin / RNA polymerase II, core complex / tRNA transcription by RNA polymerase III / RNA Polymerase II Transcription Elongation / Replacement of protamines by nucleosomes in the male pronucleus / transcription by RNA polymerase I / Formation of RNA Pol II elongation complex / CENP-A containing nucleosome / Packaging Of Telomere Ends / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / transcription-coupled nucleotide-excision repair / RNA Polymerase II Pre-transcription Events / Deposition of new CENPA-containing nucleosomes at the centromere / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / embryo implantation / DNA-directed RNA polymerase complex / telomere organization / Inhibition of DNA recombination at telomere / RNA Polymerase I Promoter Opening / Meiotic synapsis / Assembly of the ORC complex at the origin of replication / Regulation of endogenous retroelements by the Human Silencing Hub (HUSH) complex / SUMOylation of chromatin organization proteins / DNA methylation / Condensation of Prophase Chromosomes / Chromatin modifications during the maternal to zygotic transition (MZT) / SIRT1 negatively regulates rRNA expression / HCMV Late Events / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / innate immune response in mucosa / PRC2 methylates histones and DNA / Regulation of endogenous retroelements by KRAB-ZFP proteins / Defective pyroptosis 類似検索 - 分子機能 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.3 Å | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 データ登録者 データ登録者 | Naganuma, M. / Kujirai, T. / Ehara, H. / Uejima, T. / Ito, T. / Goto, M. / Aoki, M. / Henmi, M. / Miyamoto-Kohno, S. / Shirouzu, M. ...Naganuma, M. / Kujirai, T. / Ehara, H. / Uejima, T. / Ito, T. / Goto, M. / Aoki, M. / Henmi, M. / Miyamoto-Kohno, S. / Shirouzu, M. / Kurumizaka, H. / Sekine, S. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 資金援助 |  日本, 9件 日本, 9件
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 引用 引用 | ジャーナル: Sci Adv / 年: 2025 タイトル: Structural insights into promoter-proximal pausing of RNA polymerase II at +1 nucleosome. 著者: Masahiro Naganuma / Tomoya Kujirai / Haruhiko Ehara / Tamami Uejima / Tomoko Ito / Mie Goto / Mari Aoki / Masami Henmi / Sayako Miyamoto-Kohno / Mikako Shirouzu / Hitoshi Kurumizaka / Shun-Ichi Sekine / 要旨: The metazoan transcription elongation complex (EC) of RNA polymerase II (RNAPII) generally stalls between the transcription start site and the first (+1) nucleosome. This promoter-proximal pausing ...The metazoan transcription elongation complex (EC) of RNA polymerase II (RNAPII) generally stalls between the transcription start site and the first (+1) nucleosome. This promoter-proximal pausing involves negative elongation factor (NELF), 5,6-dichloro-1-β-d-ribobenzimidazole sensitivity-inducing factor (DSIF), and transcription elongation factor IIS (TFIIS) and is critical for subsequent productive transcription elongation. However, the detailed pausing mechanism and the involvement of the +1 nucleosome remain enigmatic. Here, we report cryo-electron microscopy structures of ECs stalled on nucleosomal DNA. In the absence of TFIIS, the EC is backtracked/arrested due to conflicts between NELF and the nucleosome. We identified two alternative binding modes of NELF, one of which reveals a critical contact with the downstream DNA through the conserved NELF-E basic helix. Upon binding with TFIIS, the EC progressed to the nucleosome to establish a paused EC with a partially unwrapped nucleosome. This paused EC strongly restricts EC progression further downstream. These structures illuminate the mechanism of RNAPII pausing/stalling at the +1 nucleosome. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 9j0o.cif.gz | 1.4 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb9j0o.ent.gz | 1.1 MB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 9j0o.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 9j0o_validation.pdf.gz | 1.7 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 9j0o_full_validation.pdf.gz | 1.9 MB | 表示 | |
| XML形式データ | 9j0o_validation.xml.gz | 181.8 KB | 表示 | |
| CIF形式データ | 9j0o_validation.cif.gz | 290.7 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/j0/9j0oftp://data.pdbj.org/pub/pdb/validation_reports/j0/9j0o | HTTPS FTP |
-関連構造データ
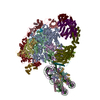
-リンク
 PDBj
PDBj





































































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-DNA-directed RNA polymerase ... , 9種, 9分子 ABCDEFGIK
| #1: タンパク質 | 分子量: 217450.078 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #2: タンパク質 | 分子量: 134041.422 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #3: タンパク質 | 分子量: 30997.557 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #4: タンパク質 | 分子量: 16331.255 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #5: タンパク質 | 分子量: 24644.318 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #6: タンパク質 | 分子量: 14477.001 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #7: タンパク質 | 分子量: 19314.283 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #9: タンパク質 | 分子量: 14541.221 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #11: タンパク質 | 分子量: 13310.284 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-DNA-directed RNA polymerases I, II, and III subunit ... , 2種, 2分子 HJ
| #8: タンパク質 | 分子量: 17162.273 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #10: タンパク質 | 分子量: 7655.123 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-タンパク質 , 5種, 9分子 Laebfcgdh
| #12: タンパク質 | 分子量: 7018.244 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) | ||||||
|---|---|---|---|---|---|---|---|
| #18: タンパク質 | 分子量: 15360.983 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: H3-3A, H3.3A, H3F3, H3F3A, PP781, H3-3B, H3.3B, H3F3B 発現宿主:  #19: タンパク質 | 分子量: 11394.426 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: HIST1H4A, H4/A, H4FA, HIST1H4B, H4/I, H4FI, HIST1H4C, H4/G, H4FG, HIST1H4D, H4/B, H4FB, HIST1H4E, H4/J, H4FJ, HIST1H4F, H4/C, H4FC, HIST1H4H, H4/H, H4FH, HIST1H4I, H4/M, H4FM, HIST1H4J, ...遺伝子: HIST1H4A, H4/A, H4FA, HIST1H4B, H4/I, H4FI, HIST1H4C, H4/G, H4FG, HIST1H4D, H4/B, H4FB, HIST1H4E, H4/J, H4FJ, HIST1H4F, H4/C, H4FC, HIST1H4H, H4/H, H4FH, HIST1H4I, H4/M, H4FM, HIST1H4J, H4/E, H4FE, HIST1H4K, H4/D, H4FD, HIST1H4L, H4/K, H4FK, HIST2H4A, H4/N, H4F2, H4FN, HIST2H4, HIST2H4B, H4/O, H4FO, HIST4H4 発現宿主: #20: タンパク質 | 分子量: 14165.551 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: HIST1H2AB, H2AFM, HIST1H2AE, H2AFA / 発現宿主: #21: タンパク質 | 分子量: 13935.239 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: HIST1H2BJ, H2BFR / 発現宿主: |
-Transcription elongation factor ... , 2種, 2分子 YZ
| #13: タンパク質 | 分子量: 13297.278 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: SUPT4H1, SPT4H, SUPT4H / 発現宿主: |
|---|---|
| #14: タンパク質 | 分子量: 123297.812 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: SUPT5H, SPT5, SPT5H / 発現宿主: |
-DNA鎖 , 2種, 2分子 NT
| #15: DNA鎖 | 分子量: 61381.906 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) |
|---|---|
| #17: DNA鎖 | 分子量: 60869.977 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) |
-RNA鎖 , 1種, 1分子 P
| #16: RNA鎖 | 分子量: 5037.968 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) |
|---|
-Negative elongation factor ... , 4種, 4分子 UWVX
| #22: タンパク質 | 分子量: 57343.598 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: NELFA, WHSC2, P/OKcl.15発現宿主:   Spodoptera frugiperda (ツマジロクサヨトウ) Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q9H3P2 |
|---|---|
| #23: タンパク質 | 分子量: 68547.758 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: NELFCD, NELFD, TH1, TH1L, HSPC130発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q8IXH7 |
| #24: タンパク質 | 分子量: 65779.227 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: NELFB, COBRA1, KIAA1182発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q8WX92 |
| #25: タンパク質 | 分子量: 45852.602 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: NELFE, RD, RDBP発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: P18615 |
-非ポリマー , 2種, 10分子 

| #26: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 9 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 9 / 由来タイプ: 合成 / 式: Zn#27: 化合物 | ChemComp-MG / |  分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | N |
|---|---|
| Has protein modification | N |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 由来(天然) |
| ||||||||||||||||||||||||||||||||||||
| 由来(組換発現) |
| ||||||||||||||||||||||||||||||||||||
| 緩衝液 | pH: 7.9 | ||||||||||||||||||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||||||||
| 急速凍結 | 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: TFS KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 最大 デフォーカス(公称値): 2500 nm / 最小 デフォーカス(公称値): 1000 nm |
| 撮影 | 電子線照射量: 57 e/Å2 / フィルム・検出器のモデル: GATAN K3 (6k x 4k) |
- 解析
解析
| EMソフトウェア | 名称: PHENIX / カテゴリ: モデル精密化 | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.3 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 24102 / 対称性のタイプ: POINT | ||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: AB INITIO MODEL | ||||||||||||||||||||||||
| 拘束条件 |
|
