ジャーナル: FEBS J / 年: 2025 タイトル: Bacterial transcriptional repressor NrdR - a flexible multifactorial nucleotide sensor. 著者: Inna Rozman Grinberg / Ornella Bimaï / Saher Shahid / Lena Philipp / Markel Martínez-Carranza / Ipsita Banerjee / Daniel Lundin / Pål Stenmark / Britt-Marie Sjöberg / Derek T Logan / 要旨: NrdR is a bacterial transcriptional repressor consisting of a zinc (Zn)-ribbon domain followed by an ATP-cone domain. Understanding its mechanism of action could aid the design of novel ...NrdR is a bacterial transcriptional repressor consisting of a zinc (Zn)-ribbon domain followed by an ATP-cone domain. Understanding its mechanism of action could aid the design of novel antibacterials. NrdR binds specifically to two "NrdR boxes" upstream of ribonucleotide reductase operons, of which Escherichia coli has three: nrdHIEF, nrdDG and nrdAB, in the last of which we identified a new box. We show that E. coli NrdR (EcoNrdR) has similar binding strength to all three sites when loaded with ATP plus deoxyadenosine triphosphate (dATP) or equivalent diphosphate combinations. No other combination of adenine nucleotides promotes binding to DNA. We present crystal structures of EcoNrdR-ATP-dATP and EcoNrdR-ADP-dATP, which are the first high-resolution crystal structures of an NrdR. We have also determined cryo-electron microscopy structures of DNA-bound EcoNrdR-ATP-dATP and novel filaments of EcoNrdR-ATP. Tetrameric forms of EcoNrdR involve alternating interactions between pairs of Zn-ribbon domains and ATP-cones. The structures reveal considerable flexibility in relative orientation of ATP-cones vs Zn-ribbon domains. The structure of DNA-bound EcoNrdR-ATP-dATP shows that significant conformational rearrangements between ATP-cones and Zn-ribbons accompany DNA binding while the ATP-cones retain the same relative orientation. In contrast, ATP-loaded EcoNrdR filaments show rearrangements of the ATP-cone pairs and sequester the DNA-binding residues of NrdR such that they are unable to bind to DNA. Our results, in combination with a previous structural and biochemical study, point to highly flexible EcoNrdR structures that, when loaded with the correct nucleotides, adapt to an optimal promoter-binding conformation.
温度: 293 K / 手法: 蒸気拡散法, シッティングドロップ法 詳細: Protein at 8 mg/ml in 1.5 mM ADP, 1.1 mM AMPPNP, 25 mM Tris-HCl pH 9 (at 277 K), 300 mM NaCl, 1 mM TCEP, 10 mM MgCl2. 150 nL protein were mixed with with 50 nL reservoir solution containing 0. ...詳細: Protein at 8 mg/ml in 1.5 mM ADP, 1.1 mM AMPPNP, 25 mM Tris-HCl pH 9 (at 277 K), 300 mM NaCl, 1 mM TCEP, 10 mM MgCl2. 150 nL protein were mixed with with 50 nL reservoir solution containing 0.1 M Bis-Tris propane pH 7.5, 0.2 M potassium thiocyanate, 20% (w/v) PEG3350 at 293 K. Crystals were harvested in paraffin oil.
-
データ収集
回折
平均測定温度: 100 K / Serial crystal experiment: N
放射光源
由来: シンクロトロン / サイト: MAX IV / ビームライン: BioMAX / 波長: 0.97963 Å
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報
 X線回折 /
X線回折 /  データ登録者
データ登録者 スウェーデン,
スウェーデン,  米国, 8件
米国, 8件  引用
引用 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード



 PDBj
PDBj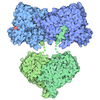











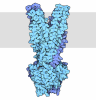
 集合体
集合体


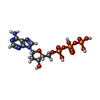



 分子量: 491.182 Da / 分子数: 8 / 由来タイプ: 合成 / 式: C10H16N5O12P3 / タイプ: SUBJECT OF INVESTIGATION
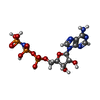
分子量: 491.182 Da / 分子数: 8 / 由来タイプ: 合成 / 式: C10H16N5O12P3 / タイプ: SUBJECT OF INVESTIGATION 分子量: 506.196 Da / 分子数: 8 / 由来タイプ: 合成 / 式: C10H17N6O12P3 / タイプ: SUBJECT OF INVESTIGATION
分子量: 506.196 Da / 分子数: 8 / 由来タイプ: 合成 / 式: C10H17N6O12P3 / タイプ: SUBJECT OF INVESTIGATION 分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn / タイプ: SUBJECT OF INVESTIGATION
分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn / タイプ: SUBJECT OF INVESTIGATION 分子量: 24.305 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION
分子量: 24.305 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION 試料調製
試料調製 解析
解析