Ministry of Education, Culture, Sports, Science and Technology (Japan)
jp15076210
Japan
Ministry of Education, Culture, Sports, Science and Technology (Japan)
jp20050030
Japan
Ministry of Education, Culture, Sports, Science and Technology (Japan)
jp22018027
Japan
Ministry of Education, Culture, Sports, Science and Technology (Japan)
jp23120525
Japan
Ministry of Education, Culture, Sports, Science and Technology (Japan)
jp25120725
Japan
Ministry of Education, Culture, Sports, Science and Technology (Japan)
jp15H01647
Japan
Ministry of Education, Culture, Sports, Science and Technology (Japan)
jp17H05891
Japan
Citation
Journal: FEBS J / Year: 2025 Title: CryoEM and crystal structure analyses reveal the indirect role played by Trp89 in glutamate dehydrogenase enzymatic reactions. Authors: Taiki Wakabayashi / Yuka Matsui / Masayoshi Nakasako / Abstract: Glutamate dehydrogenase from Thermococcus profundus is a homo-hexameric enzyme that catalyzes the reversible deamination of glutamate to 2-oxoglutarate in the presence of a cofactor. In each subunit, ...Glutamate dehydrogenase from Thermococcus profundus is a homo-hexameric enzyme that catalyzes the reversible deamination of glutamate to 2-oxoglutarate in the presence of a cofactor. In each subunit, a large active-site cleft is formed between the two functional domains, one of which displays motion to open and close the cleft. Trp89 in the cleft displays two sidechain conformers in the open cleft and a single conformer in the closed cleft. To reveal the role of the Trp89 sidechain in the domain motion, we mutated Trp89 to phenylalanine. Despite the Trp89 sidechain being located away from the reaction center, the catalytic constant decreased to 1/38-fold of that of the wild-type without a fatal reduction of the affinities to the cofactor and ligand molecules. To understand the molecular mechanism underlying this reduction, we determined the crystal structure in the unliganded state and the metastable conformations appearing in the steady stage of the reaction using cryo-electron microscopy (cryoEM). The four identified metastable conformations were similar to the three conformations observed in the wild-type, but their populations were different from those of the wild-type. In addition, a conformation with a completely closed active-site cleft necessary for the reaction to proceed was quite rare. The crystal structure and the four metastable conformations suggested that the reduction in the catalytic constant could be attributed to changes in the interactions between Gln13 and the 89th side chains, preventing the closing domain motion.
Mass: 18.015 Da / Num. of mol.: 948 / Source method: isolated from a natural source / Formula: H2O
-
Details
Has ligand of interest
N
Has protein modification
N
-
Experimental details
-
Experiment
Experiment
Method: X-RAY DIFFRACTION / Number of used crystals: 1
-
Sample preparation
Crystal
Density Matthews: 4.04 Å3/Da / Density % sol: 69.58 %
Crystal grow
Temperature: 293 K / Method: vapor diffusion, hanging drop / pH: 3.8 Details: 1.6 M lithium sulfate (Wako, Osaka, Japan), 1.7%(w/v) poly-ethylene glycol 8000 (PEG800) (Sigma-Aldrich, St. Louis, USA) and 0.1 M sodium acetate (Wako)
-
Data collection
Diffraction
Mean temperature: 100 K / Serial crystal experiment: N
 Movie
Movie Controller
Controller

 Yorodumi
Yorodumi Open data
Open data
 Basic information
Basic information Components
Components Keywords
Keywords Function and homology information
Function and homology information
 Thermococcus profundus (archaea)
Thermococcus profundus (archaea) X-RAY DIFFRACTION /
X-RAY DIFFRACTION /  Authors
Authors Japan, 13items
Japan, 13items  Citation
Citation Structure visualization
Structure visualization Downloads & links
Downloads & links Other downloads
Other downloads






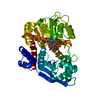


 PDBj
PDBj

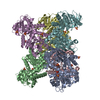
 Assembly
Assembly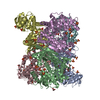

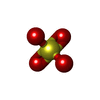
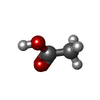


 Mass: 96.063 Da / Num. of mol.: 43 / Source method: isolated from a natural source / Formula: SO4
Mass: 96.063 Da / Num. of mol.: 43 / Source method: isolated from a natural source / Formula: SO4 Mass: 60.052 Da / Num. of mol.: 8 / Source method: isolated from a natural source / Formula: C2H4O2
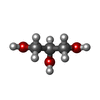
Mass: 60.052 Da / Num. of mol.: 8 / Source method: isolated from a natural source / Formula: C2H4O2 Mass: 92.094 Da / Num. of mol.: 10 / Source method: isolated from a natural source / Formula: C3H8O3
Mass: 92.094 Da / Num. of mol.: 10 / Source method: isolated from a natural source / Formula: C3H8O3 Mass: 22.990 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Formula: Na
Mass: 22.990 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Formula: Na Sample preparation
Sample preparation Processing
Processing