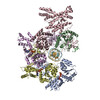
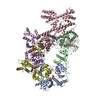
ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 8s0e | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | H. sapiens OCCM bound to double stranded DNA | |||||||||||||||
 要素 要素 |
| |||||||||||||||
 キーワード キーワード | REPLICATION / AAA+ ATPase / DNA helicase | |||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報DNA replication preinitiation complex assembly / response to sorbitol / positive regulation of chromosome segregation / positive regulation of chromatin binding / cellular response to vasopressin / polar body extrusion after meiotic divisions / CDC6 association with the ORC:origin complex / origin recognition complex / positive regulation of DNA-templated DNA replication / regulation of nuclear cell cycle DNA replication ...DNA replication preinitiation complex assembly / response to sorbitol / positive regulation of chromosome segregation / positive regulation of chromatin binding / cellular response to vasopressin / polar body extrusion after meiotic divisions / CDC6 association with the ORC:origin complex / origin recognition complex / positive regulation of DNA-templated DNA replication / regulation of nuclear cell cycle DNA replication / E2F-enabled inhibition of pre-replication complex formation / Switching of origins to a post-replicative state / Unwinding of DNA / negative regulation of DNA-templated DNA replication / nuclear origin of replication recognition complex / traversing start control point of mitotic cell cycle / Regulation of MITF-M-dependent genes involved in DNA replication, damage repair and senescence / alpha DNA polymerase:primase complex / mitotic DNA replication / DNA replication checkpoint signaling / regulation of phosphorylation / attachment of mitotic spindle microtubules to kinetochore / CMG complex / inner kinetochore / nuclear pre-replicative complex / DNA replication preinitiation complex / MCM complex / regulation of cyclin-dependent protein serine/threonine kinase activity / mitotic DNA replication checkpoint signaling / double-strand break repair via break-induced replication / mitotic DNA replication initiation / Transcription of E2F targets under negative control by DREAM complex / neural precursor cell proliferation / regulation of mitotic metaphase/anaphase transition / regulation of DNA-templated DNA replication initiation / DNA strand elongation involved in DNA replication / negative regulation of DNA replication / positive regulation of cytokinesis / spindle midzone / G1/S-Specific Transcription / negative regulation of cell cycle / regulation of DNA replication / cellular response to angiotensin / DNA replication origin binding / cochlea development / protein polymerization / Activation of the pre-replicative complex / DNA replication initiation / glial cell proliferation / Activation of ATR in response to replication stress / heterochromatin / intercellular bridge / DNA polymerase binding / cellular response to interleukin-4 / DNA helicase activity / protein serine/threonine kinase binding / Assembly of the ORC complex at the origin of replication / cellular response to epidermal growth factor stimulus / Assembly of the pre-replicative complex / positive regulation of DNA replication / CDK-mediated phosphorylation and removal of Cdc6 / helicase activity / kinetochore / Orc1 removal from chromatin / cellular response to xenobiotic stimulus / positive regulation of fibroblast proliferation / spindle pole / mitotic spindle / mitotic cell cycle / nucleosome assembly / single-stranded DNA binding / histone binding / DNA helicase / chromosome, telomeric region / DNA replication / cell population proliferation / nuclear body / cilium / negative regulation of cell population proliferation / cell division / nucleotide binding / intracellular membrane-bounded organelle / apoptotic process / DNA damage response / centrosome / chromatin binding / chromatin / nucleolus / perinuclear region of cytoplasm / enzyme binding / negative regulation of transcription by RNA polymerase II / ATP hydrolysis activity / DNA binding / zinc ion binding / nucleoplasm / ATP binding / metal ion binding / identical protein binding / nucleus / membrane 類似検索 - 分子機能 | |||||||||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト)synthetic construct (人工物) | |||||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.8 Å | |||||||||||||||
 データ登録者 データ登録者 | Greiwe, J.F. / Weissmann, F. / Diffley, J.F.X. / Costa, A. | |||||||||||||||
| 資金援助 |  英国, European Union, 4件 英国, European Union, 4件
| |||||||||||||||
 引用 引用 | ジャーナル: Nature / 年: 2024 タイトル: MCM double hexamer loading visualized with human proteins. 著者: Florian Weissmann / Julia F Greiwe / Thomas Pühringer / Evelyn L Eastwood / Emma C Couves / Thomas C R Miller / John F X Diffley / Alessandro Costa /  要旨: Eukaryotic DNA replication begins with the loading of the MCM replicative DNA helicase as a head-to-head double hexamer at origins of DNA replication. Our current understanding of how the double ...Eukaryotic DNA replication begins with the loading of the MCM replicative DNA helicase as a head-to-head double hexamer at origins of DNA replication. Our current understanding of how the double hexamer is assembled by the origin recognition complex (ORC), CDC6 and CDT1 comes mostly from budding yeast. Here we characterize human double hexamer (hDH) loading using biochemical reconstitution and cryo-electron microscopy with purified proteins. We show that the human double hexamer engages DNA differently from the yeast double hexamer (yDH), and generates approximately five base pairs of underwound DNA at the interface between hexamers, as seen in hDH isolated from cells. We identify several differences from the yeast double hexamer in the order of factor recruitment and dependencies during hDH assembly. Unlike in yeast, the ORC6 subunit of the ORC is not essential for initial MCM recruitment or hDH loading, but contributes to an alternative hDH assembly pathway that requires an intrinsically disordered region in ORC1, which may work through a MCM-ORC intermediate. Our work presents a detailed view of how double hexamers are assembled in an organism that uses sequence-independent replication origins, provides further evidence for diversity in eukaryotic double hexamer assembly mechanisms, and represents a first step towards reconstitution of DNA replication initiation with purified human proteins. #1: ジャーナル: Biorxiv / 年: 2024タイトル: MCM Double Hexamer Loading Visualised with Human Proteins 著者: Weissmann, F. / Greiwe, J.F. / Puhringer, T. / Miller, T.C.R. / Diffley, J.F.X. / Costa, A. #2: ジャーナル: Acta Crystallogr D Struct Biol / 年: 2019 タイトル: Macromolecular structure determination using X-rays, neutrons and electrons: recent developments in Phenix. 著者: Dorothee Liebschner / Pavel V Afonine / Matthew L Baker / Gábor Bunkóczi / Vincent B Chen / Tristan I Croll / Bradley Hintze / Li Wei Hung / Swati Jain / Airlie J McCoy / Nigel W Moriarty / ...著者: Dorothee Liebschner / Pavel V Afonine / Matthew L Baker / Gábor Bunkóczi / Vincent B Chen / Tristan I Croll / Bradley Hintze / Li Wei Hung / Swati Jain / Airlie J McCoy / Nigel W Moriarty / Robert D Oeffner / Billy K Poon / Michael G Prisant / Randy J Read / Jane S Richardson / David C Richardson / Massimo D Sammito / Oleg V Sobolev / Duncan H Stockwell / Thomas C Terwilliger / Alexandre G Urzhumtsev / Lizbeth L Videau / Christopher J Williams / Paul D Adams /   要旨: Diffraction (X-ray, neutron and electron) and electron cryo-microscopy are powerful methods to determine three-dimensional macromolecular structures, which are required to understand biological ...Diffraction (X-ray, neutron and electron) and electron cryo-microscopy are powerful methods to determine three-dimensional macromolecular structures, which are required to understand biological processes and to develop new therapeutics against diseases. The overall structure-solution workflow is similar for these techniques, but nuances exist because the properties of the reduced experimental data are different. Software tools for structure determination should therefore be tailored for each method. Phenix is a comprehensive software package for macromolecular structure determination that handles data from any of these techniques. Tasks performed with Phenix include data-quality assessment, map improvement, model building, the validation/rebuilding/refinement cycle and deposition. Each tool caters to the type of experimental data. The design of Phenix emphasizes the automation of procedures, where possible, to minimize repetitive and time-consuming manual tasks, while default parameters are chosen to encourage best practice. A graphical user interface provides access to many command-line features of Phenix and streamlines the transition between programs, project tracking and re-running of previous tasks. | |||||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 8s0e.cif.gz | 1.2 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb8s0e.ent.gz | 822.7 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 8s0e.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 8s0e_validation.pdf.gz | 2.1 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 8s0e_full_validation.pdf.gz | 2.1 MB | 表示 | |
| XML形式データ | 8s0e_validation.xml.gz | 146.6 KB | 表示 | |
| CIF形式データ | 8s0e_validation.cif.gz | 221.3 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/s0/8s0eftp://data.pdbj.org/pub/pdb/validation_reports/s0/8s0e | HTTPS FTP |
-関連構造データ













-リンク
 PDBj
PDBj








































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-DNA鎖 , 2種, 2分子 XY
| #1: DNA鎖 | 分子量: 12000.695 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) synthetic construct (人工物) |
|---|---|
| #2: DNA鎖 | 分子量: 12009.710 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) synthetic construct (人工物) |
-DNA replication licensing factor ... , 6種, 6分子 524673
| #3: タンパク質 | 分子量: 82406.633 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: MCM5, CDC46発現宿主:   Spodoptera frugiperda (ツマジロクサヨトウ) Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: P33992, DNA helicase |
|---|---|
| #5: タンパク質 | 分子量: 102034.102 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: MCM2, BM28, CCNL1, CDCL1, KIAA0030発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: P49736, DNA helicase |
| #6: タンパク質 | 分子量: 96702.891 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: MCM4, CDC21発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: P33991, DNA helicase |
| #7: タンパク質 | 分子量: 93010.273 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: MCM6発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q14566, DNA helicase |
| #8: タンパク質 | 分子量: 81411.875 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: MCM7, CDC47, MCM2発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: P33993, DNA helicase |
| #12: タンパク質 | 分子量: 91297.023 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: MCM3発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: P25205, DNA helicase |
-タンパク質 , 2種, 2分子 8G
| #4: タンパク質 | 分子量: 60483.371 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: CDT1発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q9H211 |
|---|---|
| #11: タンパク質 | 分子量: 62820.355 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: CDC6, CDC18L発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q99741 |
-Origin recognition complex subunit ... , 5種, 5分子 BCADE
| #9: タンパク質 | 分子量: 66063.375 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: ORC2, ORC2L発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q13416 |
|---|---|
| #10: タンパク質 | 分子量: 82365.055 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: ORC3, LATHEO, ORC3L発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q9UBD5 |
| #13: タンパク質 | 分子量: 97499.867 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: ORC1, ORC1L, PARC1発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q13415 |
| #14: タンパク質 | 分子量: 50443.266 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: ORC4, ORC4L発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: O43929 |
| #15: タンパク質 | 分子量: 50349.934 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: ORC5, ORC5L発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: O43913 |
-非ポリマー , 3種, 11分子 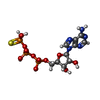


| #16: 化合物 | ChemComp-AGS /  分子量: 523.247 Da / 分子数: 5 / 由来タイプ: 合成 / 式: C10H16N5O12P3S / タイプ: SUBJECT OF INVESTIGATION 分子量: 523.247 Da / 分子数: 5 / 由来タイプ: 合成 / 式: C10H16N5O12P3S / タイプ: SUBJECT OF INVESTIGATIONコメント: ATP-gamma-S, エネルギー貯蔵分子類似体*YM #17: 化合物 | ChemComp-MG /  分子量: 24.305 Da / 分子数: 5 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION 分子量: 24.305 Da / 分子数: 5 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION#18: 化合物 | ChemComp-ZN / |  分子量: 65.409 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Zn / タイプ: SUBJECT OF INVESTIGATION 分子量: 65.409 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Zn / タイプ: SUBJECT OF INVESTIGATION |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | Y |
|---|---|
| Has protein modification | N |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 値: 1.1 MDa / 実験値: NO | ||||||||||||||||||||||||
| 由来(天然) |
| ||||||||||||||||||||||||
| 由来(組換発現) |
| ||||||||||||||||||||||||
| 緩衝液 | pH: 7.5 | ||||||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES 詳細: The MCM recruitment reaction was reconstituted in vitro using purified H. sapiens proteins, a short DNA template and ATPgammaS. Four microlitres of the entire reaction was applied on a grid ...詳細: The MCM recruitment reaction was reconstituted in vitro using purified H. sapiens proteins, a short DNA template and ATPgammaS. Four microlitres of the entire reaction was applied on a grid and incubated for 1 min at room temperature before blotting with filter paper for 5 s and plunge-freezing in liquid ethane. | ||||||||||||||||||||||||
| 試料支持 | グリッドの材料: GOLD / グリッドのサイズ: 300 divisions/in. / グリッドのタイプ: UltrAuFoil R1.2/1.3 | ||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 90 % / 凍結前の試料温度: 295 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 130000 X / 最大 デフォーカス(公称値): 2500 nm / 最小 デフォーカス(公称値): 1000 nm |
| 試料ホルダ | 凍結剤: NITROGEN |
| 撮影 | 平均露光時間: 9.4 sec. / 電子線照射量: 49.28 e/Å2 / 検出モード: COUNTING フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 撮影したグリッド数: 1 / 実像数: 31569 |
| 電子光学装置 | エネルギーフィルター名称: GIF Bioquantum |
- 解析
解析
| EMソフトウェア | 名称: PHENIX / バージョン: 1.21_5207 / カテゴリ: モデル精密化 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.8 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 34116 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | 3D fitting-ID: 1
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 | 交差検証法: NONE 立体化学のターゲット値: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 289.14 Å2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
|