 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8ptk | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
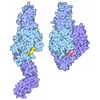
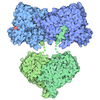
| Title | Composite structure of Dynein-Dynactin-JIP3-LIS1 | ||||||||||||
 Components Components |
| ||||||||||||
 Keywords Keywords | MOTOR PROTEIN / Dynein / AAA-Atpase / p150 / LIS1 / Dynactin / JIP3 | ||||||||||||
| Function / homology |  Function and homology information Function and homology informationintracellular transport of viral protein in host cell / nitric-oxide synthase inhibitor activity / ameboidal-type cell migration / microtubule cytoskeleton organization involved in establishment of planar polarity / establishment of planar polarity of embryonic epithelium / retrograde axonal transport of mitochondrion / Regulation of actin dynamics for phagocytic cup formation / EPHB-mediated forward signaling / Adherens junctions interactions / VEGFA-VEGFR2 Pathway ...intracellular transport of viral protein in host cell / nitric-oxide synthase inhibitor activity / ameboidal-type cell migration / microtubule cytoskeleton organization involved in establishment of planar polarity / establishment of planar polarity of embryonic epithelium / retrograde axonal transport of mitochondrion / Regulation of actin dynamics for phagocytic cup formation / EPHB-mediated forward signaling / Adherens junctions interactions / VEGFA-VEGFR2 Pathway / Cell-extracellular matrix interactions / RHO GTPases Activate WASPs and WAVEs / MAP2K and MAPK activation / Formation of the canonical BAF (cBAF) complex / Formation of the polybromo-BAF (pBAF) complex / Formation of the embryonic stem cell BAF (esBAF) complex / Formation of the non-canonical BAF (ncBAF) complex / UCH proteinases / negative regulation of DNA strand resection involved in replication fork processing / 1-alkyl-2-acetylglycerophosphocholine esterase complex / RHOF GTPase cycle / Regulation of CDH1 Function / Formation of the dystrophin-glycoprotein complex (DGC) / Gap junction degradation / Formation of annular gap junctions / secretory vesicle / Clathrin-mediated endocytosis / dynactin complex / centriolar subdistal appendage / corpus callosum morphogenesis / negative regulation of phosphorylation / maintenance of centrosome location / centriole-centriole cohesion / visual behavior / intraciliary retrograde transport / platelet activating factor metabolic process / positive regulation of neuromuscular junction development / radial glia-guided pyramidal neuron migration / Regulation of PLK1 Activity at G2/M Transition / Loss of Nlp from mitotic centrosomes / Loss of proteins required for interphase microtubule organization from the centrosome / Anchoring of the basal body to the plasma membrane / AURKA Activation by TPX2 / microtubule anchoring at centrosome / establishment of centrosome localization / acrosome assembly / cerebral cortex neuron differentiation / Recruitment of mitotic centrosome proteins and complexes / nuclear membrane disassembly / central region of growth cone / F-actin capping protein complex / WASH complex / anterograde axonal protein transport / microtubule sliding / dynein light chain binding / transport along microtubule / ventral spinal cord development / dynein heavy chain binding / motile cilium assembly / Activation of BIM and translocation to mitochondria / positive regulation of cytokine-mediated signaling pathway / microtubule organizing center organization / positive regulation of embryonic development / layer formation in cerebral cortex / retromer complex / cytoskeleton-dependent cytokinesis / interneuron migration / dynein complex / astral microtubule / auditory receptor cell development / microtubule plus-end / cortical microtubule organization / cellular response to cytochalasin B / Intraflagellar transport / positive regulation of microtubule nucleation / myeloid leukocyte migration / MAP kinase scaffold activity / reelin-mediated signaling pathway / positive regulation of intracellular transport / regulation of transepithelial transport / regulation of metaphase plate congression / positive regulation of spindle assembly / JUN kinase binding / positive regulation of dendritic spine morphogenesis / morphogenesis of a polarized epithelium / structural constituent of postsynaptic actin cytoskeleton / melanosome transport / osteoclast development / protein localization to adherens junction / negative regulation of nitric oxide biosynthetic process / establishment of spindle localization / regulation of G protein-coupled receptor signaling pathway / barbed-end actin filament capping / dense body / stereocilium / Neutrophil degranulation / Tat protein binding / microtubule plus-end binding / brain morphogenesis / coronary vasculature development Similarity search - Function | ||||||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 10 Å | ||||||||||||
 Authors Authors | Singh, K. / Lau, C.K. / Manigrasso, G. / Gassmann, R. / Carter, A.P. | ||||||||||||
| Funding support |  United Kingdom, European Union, 3items United Kingdom, European Union, 3items
| ||||||||||||
 Citation Citation | Journal: Science / Year: 2024 Title: Molecular mechanism of dynein-dynactin complex assembly by LIS1. Authors: Kashish Singh / Clinton K Lau / Giulia Manigrasso / José B Gama / Reto Gassmann / Andrew P Carter /  Abstract: Cytoplasmic dynein is a microtubule motor vital for cellular organization and division. It functions as a ~4-megadalton complex containing its cofactor dynactin and a cargo-specific coiled-coil ...Cytoplasmic dynein is a microtubule motor vital for cellular organization and division. It functions as a ~4-megadalton complex containing its cofactor dynactin and a cargo-specific coiled-coil adaptor. However, how dynein and dynactin recognize diverse adaptors, how they interact with each other during complex formation, and the role of critical regulators such as lissencephaly-1 (LIS1) protein (LIS1) remain unclear. In this study, we determined the cryo-electron microscopy structure of dynein-dynactin on microtubules with LIS1 and the lysosomal adaptor JIP3. This structure reveals the molecular basis of interactions occurring during dynein activation. We show how JIP3 activates dynein despite its atypical architecture. Unexpectedly, LIS1 binds dynactin's p150 subunit, tethering it along the length of dynein. Our data suggest that LIS1 and p150 constrain dynein-dynactin to ensure efficient complex formation. | ||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8ptk.cif.gz | 4.4 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8ptk.ent.gz | Display | PDB format | |
| PDBx/mmJSON format | 8ptk.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/pt/8ptkftp://data.pdbj.org/pub/pdb/validation_reports/pt/8ptk | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  17873MC  8pqvC  8pqwC  8pqyC 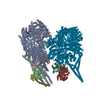 8pqzC  8pr0C  8pr1C  8pr2C  8pr3C  8pr4C  8pr5C M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |











-Links
 PDBj
PDBj


























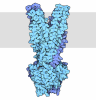

- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-Protein , 8 types, 19 molecules 1234ABCDEFGIHJKLUXx
| #1: Protein | Mass: 46709.984 Da / Num. of mol.: 4 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: PAFAH1B1, LIS1, MDCR, MDS, PAFAHA / Production host:   Spodoptera frugiperda (fall armyworm) / References: UniProt: P43034 Spodoptera frugiperda (fall armyworm) / References: UniProt: P43034#2: Protein | Mass: 42670.688 Da / Num. of mol.: 8 / Source method: isolated from a natural source / Source: (natural) #3: Protein | | Mass: 41782.660 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) #4: Protein | | Mass: 46250.785 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) #5: Protein | | Mass: 33059.848 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) #6: Protein | | Mass: 30669.768 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) #10: Protein | | Mass: 20703.910 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) #12: Protein | Mass: 65975.398 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: MAPK8IP3, JIP3, KIAA1066 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: Q9UPT6 |
|---|
-Dynactin subunit ... , 5 types, 10 molecules MNPQORSTWY
| #7: Protein | Mass: 44704.414 Da / Num. of mol.: 4 / Source method: isolated from a natural source / Source: (natural) #8: Protein | Mass: 21192.477 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #9: Protein | Mass: 142015.484 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #11: Protein | | Mass: 20150.533 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) #13: Protein | | Mass: 52920.434 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|
-Dynein light chain ... , 3 types, 12 molecules abdiklvystwz
| #14: Protein | Mass: 10381.899 Da / Num. of mol.: 4 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: DYNLL1, DLC1, DNCL1, DNCLC1, HDLC1 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: P63167#18: Protein | Mass: 12461.996 Da / Num. of mol.: 4 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: DYNLT1 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: P63172#19: Protein | Mass: 10934.576 Da / Num. of mol.: 4 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: DYNLRB1, BITH, DNCL2A, DNLC2A, ROBLD1, HSPC162 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: Q9NP97 |
|---|
-Cytoplasmic dynein 1 ... , 3 types, 12 molecules efmnghopjqru
| #15: Protein | Mass: 533055.125 Da / Num. of mol.: 4 / Mutation: R1567E, K1610E Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: DYNC1H1, DHC1, DNCH1, DNCL, DNECL, DYHC, KIAA0325 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: Q14204#16: Protein | Mass: 68442.141 Da / Num. of mol.: 4 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: DYNC1I2, DNCI2, DNCIC2 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: Q13409#17: Protein | Mass: 54173.156 Da / Num. of mol.: 4 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: DYNC1LI2, DNCLI2, LIC2 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: O43237 |
|---|
-Non-polymers , 5 types, 35 molecules 



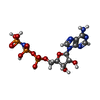
| #20: Chemical | ChemComp-ADP /  Mass: 427.201 Da / Num. of mol.: 16 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Feature type: SUBJECT OF INVESTIGATION / Comment: ADP, energy-carrying molecule*YM Mass: 427.201 Da / Num. of mol.: 16 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Feature type: SUBJECT OF INVESTIGATION / Comment: ADP, energy-carrying molecule*YM#21: Chemical | ChemComp-ATP /  Mass: 507.181 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Feature type: SUBJECT OF INVESTIGATION / Comment: ATP, energy-carrying molecule*YM Mass: 507.181 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Feature type: SUBJECT OF INVESTIGATION / Comment: ATP, energy-carrying molecule*YM#22: Chemical |  Mass: 65.409 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: Zn Mass: 65.409 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: Zn#23: Chemical | ChemComp-MG /  Mass: 24.305 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: Mg#24: Chemical | ChemComp-ANP /  Mass: 506.196 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: C10H17N6O12P3 / Feature type: SUBJECT OF INVESTIGATION / Comment: AMP-PNP, energy-carrying molecule analogue*YM Mass: 506.196 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: C10H17N6O12P3 / Feature type: SUBJECT OF INVESTIGATION / Comment: AMP-PNP, energy-carrying molecule analogue*YM |
|---|
-Details
| Has ligand of interest | Y |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Source (natural) |
| ||||||||||||||||||||||||
| Source (recombinant) | Organism: Spodoptera frugiperda (fall armyworm) | ||||||||||||||||||||||||
| Buffer solution | pH: 7.2 | ||||||||||||||||||||||||
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||||||
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal defocus max: 4000 nm / Nominal defocus min: 500 nm |
| Image recording | Electron dose: 53 e/Å2 / Film or detector model: GATAN K3 BIOQUANTUM (6k x 4k) |
- Processing
Processing
| EM software |
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||
| 3D reconstruction | Resolution: 10 Å / Resolution method: OTHER / Num. of particles: 700290 / Symmetry type: POINT | ||||||||||||
| Atomic model building | PDB-ID: 7Z8G Accession code: 7Z8G / Source name: PDB / Type: experimental model |

