 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 8eqt | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | Structure of SARS-CoV-2 Orf3a in plasma membrane-like environment, MSP1D1 nanodisc | ||||||
 要素 要素 | ORF3a protein | ||||||
 キーワード キーワード | VIRAL PROTEIN / Membrane protein / SARS-CoV-2 | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報host cell lysosome / symbiont-mediated activation of host reticulophagy / Maturation of protein 3a / SARS-CoV-2 modulates autophagy / : / voltage-gated calcium channel complex / host cell endoplasmic reticulum / monoatomic ion channel activity / SARS-CoV-2 targets host intracellular signalling and regulatory pathways / voltage-gated potassium channel complex ...host cell lysosome / symbiont-mediated activation of host reticulophagy / Maturation of protein 3a / SARS-CoV-2 modulates autophagy / : / voltage-gated calcium channel complex / host cell endoplasmic reticulum / monoatomic ion channel activity / SARS-CoV-2 targets host intracellular signalling and regulatory pathways / voltage-gated potassium channel complex / molecular function activator activity / cytoplasmic side of plasma membrane / Translation of Structural Proteins / Virion Assembly and Release / host cell endosome / Induction of Cell-Cell Fusion / Attachment and Entry / host cell endoplasmic reticulum membrane / host cell plasma membrane / SARS-CoV-2 activates/modulates innate and adaptive immune responses / virion membrane / extracellular region / identical protein binding / plasma membrane 類似検索 - 分子機能 | ||||||
| 生物種 |   Severe acute respiratory syndrome coronavirus 2 (ウイルス) Severe acute respiratory syndrome coronavirus 2 (ウイルス) | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.4 Å | ||||||
 データ登録者 データ登録者 | Miller, A.N. / Houlihan, P.R. / Matamala, E. / Cabezas-Bratesco, D. / Lee, G.Y. / Cristofori-Armstrong, B. / Dilan, T.L. / Sanchez-Martinez, S. / Matthies, D. / Yan, R. ...Miller, A.N. / Houlihan, P.R. / Matamala, E. / Cabezas-Bratesco, D. / Lee, G.Y. / Cristofori-Armstrong, B. / Dilan, T.L. / Sanchez-Martinez, S. / Matthies, D. / Yan, R. / Yu, Z. / Ren, D. / Brauchi, S.E. / Clapham, D.E. | ||||||
| 資金援助 |  米国, 1件 米国, 1件
| ||||||
 引用 引用 | ジャーナル: Elife / 年: 2023 タイトル: The SARS-CoV-2 accessory protein Orf3a is not an ion channel, but does interact with trafficking proteins. 著者: Alexandria N Miller / Patrick R Houlihan / Ella Matamala / Deny Cabezas-Bratesco / Gi Young Lee / Ben Cristofori-Armstrong / Tanya L Dilan / Silvia Sanchez-Martinez / Doreen Matthies / Rui ...著者: Alexandria N Miller / Patrick R Houlihan / Ella Matamala / Deny Cabezas-Bratesco / Gi Young Lee / Ben Cristofori-Armstrong / Tanya L Dilan / Silvia Sanchez-Martinez / Doreen Matthies / Rui Yan / Zhiheng Yu / Dejian Ren / Sebastian E Brauchi / David E Clapham /  要旨: The severe acute respiratory syndrome associated coronavirus 2 (SARS-CoV-2) and SARS-CoV-1 accessory protein Orf3a colocalizes with markers of the plasma membrane, endocytic pathway, and Golgi ...The severe acute respiratory syndrome associated coronavirus 2 (SARS-CoV-2) and SARS-CoV-1 accessory protein Orf3a colocalizes with markers of the plasma membrane, endocytic pathway, and Golgi apparatus. Some reports have led to annotation of both Orf3a proteins as viroporins. Here, we show that neither SARS-CoV-2 nor SARS-CoV-1 Orf3a form functional ion conducting pores and that the conductances measured are common contaminants in overexpression and with high levels of protein in reconstitution studies. Cryo-EM structures of both SARS-CoV-2 and SARS-CoV-1 Orf3a display a narrow constriction and the presence of a positively charged aqueous vestibule, which would not favor cation permeation. We observe enrichment of the late endosomal marker Rab7 upon SARS-CoV-2 Orf3a overexpression, and co-immunoprecipitation with VPS39. Interestingly, SARS-CoV-1 Orf3a does not cause the same cellular phenotype as SARS-CoV-2 Orf3a and does not interact with VPS39. To explain this difference, we find that a divergent, unstructured loop of SARS-CoV-2 Orf3a facilitates its binding with VPS39, a HOPS complex tethering protein involved in late endosome and autophagosome fusion with lysosomes. We suggest that the added loop enhances SARS-CoV-2 Orf3a's ability to co-opt host cellular trafficking mechanisms for viral exit or host immune evasion. #1: ジャーナル: bioRxiv / 年: 2022 タイトル: The SARS-CoV-2 accessory protein Orf3a is not an ion channel, but does interact with trafficking proteins. 著者: Alexandria N Miller / Patrick R Houlihan / Ella Matamala / Deny Cabezas-Bratesco / Gi Young Lee / Ben Cristofori-Armstrong / Tanya L Dilan / Silvia Sanchez-Martinez / Doreen Matthies / Rui ...著者: Alexandria N Miller / Patrick R Houlihan / Ella Matamala / Deny Cabezas-Bratesco / Gi Young Lee / Ben Cristofori-Armstrong / Tanya L Dilan / Silvia Sanchez-Martinez / Doreen Matthies / Rui Yan / Zhiheng Yu / Dejian Ren / Sebastian E Brauchi / David E Clapham 要旨: The severe acute respiratory syndrome associated coronavirus 2 (SARS-CoV-2) and SARS-CoV-1 accessory protein Orf3a colocalizes with markers of the plasma membrane, endocytic pathway, and Golgi ...The severe acute respiratory syndrome associated coronavirus 2 (SARS-CoV-2) and SARS-CoV-1 accessory protein Orf3a colocalizes with markers of the plasma membrane, endocytic pathway, and Golgi apparatus. Some reports have led to annotation of both Orf3a proteins as a viroporin. Here we show that neither SARS-CoV-2 nor SARS-CoV-1 form functional ion conducting pores and that the conductances measured are common contaminants in overexpression and with high levels of protein in reconstitution studies. Cryo-EM structures of both SARS-CoV-2 and SARS-CoV-1 Orf3a display a narrow constriction and the presence of a basic aqueous vestibule, which would not favor cation permeation. We observe enrichment of the late endosomal marker Rab7 upon SARS-CoV-2 Orf3a overexpression, and co-immunoprecipitation with VPS39. Interestingly, SARS-CoV-1 Orf3a does not cause the same cellular phenotype as SARS-CoV-2 Orf3a and does not interact with VPS39. To explain this difference, we find that a divergent, unstructured loop of SARS-CoV-2 Orf3a facilitates its binding with VPS39, a HOPS complex tethering protein involved in late endosome and autophagosome fusion with lysosomes. We suggest that the added loop enhances SARS-CoV-2 Orf3a ability to co-opt host cellular trafficking mechanisms for viral exit or host immune evasion. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 8eqt.cif.gz | 88 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb8eqt.ent.gz | 62.7 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 8eqt.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/eq/8eqtftp://data.pdbj.org/pub/pdb/validation_reports/eq/8eqt | HTTPS FTP |
|---|
-関連構造データ







-リンク
 PDBj
PDBj
- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
| #1: タンパク質 | 分子量: 36489.445 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Severe acute respiratory syndrome coronavirus 2 (ウイルス)遺伝子: 3a / 発現宿主:  Homo sapiens (ヒト) / 参照: UniProt: P0DTC3 Homo sapiens (ヒト) / 参照: UniProt: P0DTC3#2: 化合物 | ChemComp-PEE / 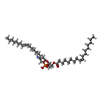  分子量: 744.034 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C41H78NO8P / コメント: DOPE, リン脂質*YM 分子量: 744.034 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C41H78NO8P / コメント: DOPE, リン脂質*YM研究の焦点であるリガンドがあるか | N | |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Structure of SARS-CoV2 Orf3a in plasma membrane-like environment, MSP1D1 nanodisc タイプ: COMPLEX / Entity ID: #1 / 由来: RECOMBINANT |
|---|---|
| 分子量 | 実験値: NO |
| 由来(天然) | 生物種: Severe acute respiratory syndrome coronavirus 2 (ウイルス) |
| 由来(組換発現) | 生物種: Homo sapiens (ヒト) |
| 緩衝液 | pH: 7.5 |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 最大 デフォーカス(公称値): 2000 nm / 最小 デフォーカス(公称値): 800 nm |
| 撮影 | 電子線照射量: 50 e/Å2 / フィルム・検出器のモデル: GATAN K3 (6k x 4k) / 撮影したグリッド数: 1 / 実像数: 15637 |
| 電子光学装置 | エネルギーフィルター名称: GIF Bioquantum |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: NONE | ||||||||||||||||||||
| 対称性 | 点対称性: C2 (2回回転対称) | ||||||||||||||||||||
| 3次元再構成 | 解像度: 3.4 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 125625 / クラス平均像の数: 1 / 対称性のタイプ: POINT | ||||||||||||||||||||
| 原子モデル構築 | プロトコル: AB INITIO MODEL |
