National Institutes of Health/National Heart, Lung, and Blood Institute (NIH/NHLBI)
R01HL036153
米国
引用
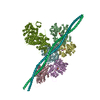
ジャーナル: J Gen Physiol / 年: 2023 タイトル: Myosin loop-4 is critical for optimal tropomyosin repositioning on actin during muscle activation and relaxation. 著者: Matthew H Doran / Michael J Rynkiewicz / Elumalai Pavadai / Skylar M L Bodt / David Rasicci / Jeffrey R Moore / Christopher M Yengo / Esther Bullitt / William Lehman / 要旨: During force-generating steps of the muscle crossbridge cycle, the tip of the myosin motor, specifically loop-4, contacts the tropomyosin cable of actin filaments. In the current study, we determined ...During force-generating steps of the muscle crossbridge cycle, the tip of the myosin motor, specifically loop-4, contacts the tropomyosin cable of actin filaments. In the current study, we determined the corresponding effect of myosin loop-4 on the regulatory positioning of tropomyosin on actin. To accomplish this, we compared high-resolution cryo-EM structures of myosin S1-decorated thin filaments containing either wild-type or a loop-4 mutant construct, where the seven-residue portion of myosin loop-4 that contacts tropomyosin was replaced by glycine residues, thus removing polar side chains from residues 366-372. Cryo-EM analysis of fully decorated actin-tropomyosin filaments with wild-type and mutant S1, yielded 3.4-3.6 Å resolution reconstructions, with even higher definition at the actin-myosin interface. Loop-4 densities both in wild-type and mutant S1 were clearly identified, and side chains were resolved in the wild-type structure. Aside from loop-4, actin and myosin structural domains were indistinguishable from each other when filaments were decorated with either mutant or wild-type S1. In marked contrast, the position of tropomyosin on actin in the two reconstructions differed by 3 to 4 Å. In maps of filaments containing the mutant, tropomyosin was located closer to the myosin-head and thus moved in the direction of the C-state conformation adopted by myosin-free thin filaments. Complementary interaction energy measurements showed that tropomyosin in the mutant thin filaments sits on actin in a local energy minimum, whereas tropomyosin is positioned by wild-type S1 in an energetically unfavorable location. We propose that the high potential energy associated with tropomyosin positioning in wild-type filaments favors an effective transition to B- and C-states following release of myosin from the thin filaments during relaxation.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト)
 データ登録者
データ登録者 米国, 1件
米国, 1件  引用
引用 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード

 PDBj
PDBj









 集合体
集合体


 分子量: 427.201 Da / 分子数: 5 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / コメント: ADP, エネルギー貯蔵分子*YM
分子量: 427.201 Da / 分子数: 5 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / コメント: ADP, エネルギー貯蔵分子*YM
 分子量: 24.305 Da / 分子数: 5 / 由来タイプ: 合成 / 式: Mg
分子量: 24.305 Da / 分子数: 5 / 由来タイプ: 合成 / 式: Mg 試料調製
試料調製 電子顕微鏡撮影
電子顕微鏡撮影
 FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM
FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM 解析
解析
