 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 8a8f | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Crystal structure of Glc7 phosphatase in complex with the regulatory region of Ref2 | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード | GENE REGULATION / Ser/Thr phosphatase / regulatory subunit / RNA / transcription termination | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報regulation of cell budding / regulation of protein localization to cell division site involved in cytokinesis / cellular bud neck septin collar / mating projection base / positive regulation of mitotic actomyosin contractile ring assembly / regulation of cellular response to glucose starvation / Resolution of Sister Chromatid Cohesion / positive regulation of clathrin-dependent endocytosis / termination of RNA polymerase II transcription, poly(A)-coupled / sno(s)RNA 3'-end processing ...regulation of cell budding / regulation of protein localization to cell division site involved in cytokinesis / cellular bud neck septin collar / mating projection base / positive regulation of mitotic actomyosin contractile ring assembly / regulation of cellular response to glucose starvation / Resolution of Sister Chromatid Cohesion / positive regulation of clathrin-dependent endocytosis / termination of RNA polymerase II transcription, poly(A)-coupled / sno(s)RNA 3'-end processing / regulation of glycogen biosynthetic process / chitin biosynthetic process / cellular bud site selection / regulation of glycogen metabolic process / ascospore formation / positive regulation of exit from mitosis / protein phosphatase type 1 complex / incipient cellular bud site / intracellular monoatomic ion homeostasis / mRNA cleavage and polyadenylation specificity factor complex / negative regulation of actin filament polymerization / DNA replication checkpoint signaling / cellular bud neck / protein localization to kinetochore / spindle pole body / mRNA 3'-end processing / glycogen metabolic process / cell division site / protein-serine/threonine phosphatase / mitotic spindle assembly checkpoint signaling / protein serine/threonine phosphatase activity / replication fork processing / response to unfolded protein / chromosome organization / telomere maintenance / regulation of mitotic cell cycle / DNA damage checkpoint signaling / meiotic cell cycle / chromosome segregation / kinetochore / mRNA processing / regulation of cell shape / response to heat / regulation of cell cycle / chromatin binding / nucleolus / positive regulation of transcription by RNA polymerase II / RNA binding / metal ion binding / nucleus / cytoplasm / cytosol 類似検索 - 分子機能 | |||||||||
| 生物種 |  | |||||||||
| 手法 |  X線回折 / シンクロトロン / 分子置換 / 解像度: 1.85 Å X線回折 / シンクロトロン / 分子置換 / 解像度: 1.85 Å | |||||||||
 データ登録者 データ登録者 | Carminati, M. / Manav, C.M. / Bellini, D. / Passmore, L.A. | |||||||||
| 資金援助 | European Union, 2件
| |||||||||
 引用 引用 | ジャーナル: Mol Cell / 年: 2023 タイトル: A direct interaction between CPF and RNA Pol II links RNA 3' end processing to transcription. 著者: Manuel Carminati / Juan B Rodríguez-Molina / M Cemre Manav / Dom Bellini / Lori A Passmore /  要旨: Transcription termination by RNA polymerase II (RNA Pol II) is linked to RNA 3' end processing by the cleavage and polyadenylation factor (CPF or CPSF). CPF contains endonuclease, poly(A) polymerase, ...Transcription termination by RNA polymerase II (RNA Pol II) is linked to RNA 3' end processing by the cleavage and polyadenylation factor (CPF or CPSF). CPF contains endonuclease, poly(A) polymerase, and protein phosphatase activities, which cleave and polyadenylate pre-mRNAs and dephosphorylate RNA Pol II to control transcription. Exactly how the RNA 3' end processing machinery is coupled to transcription remains unclear. Here, we combine in vitro reconstitution, structural studies, and genome-wide analyses to show that yeast CPF physically and functionally interacts with RNA Pol II. Surprisingly, CPF-mediated dephosphorylation promotes the formation of an RNA Pol II stalk-to-stalk homodimer in vitro. This dimer is compatible with transcription but not with the binding of transcription elongation factors. Disruption of the dimerization interface in cells causes transcription defects, including altered RNA Pol II abundance on protein-coding genes, tRNA genes, and intergenic regions. We hypothesize that RNA Pol II dimerization may provide a mechanistic basis for the allosteric model of transcription termination. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 8a8f.cif.gz | 90.6 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb8a8f.ent.gz | 66.2 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 8a8f.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 8a8f_validation.pdf.gz | 457.7 KB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 8a8f_full_validation.pdf.gz | 460.5 KB | 表示 | |
| XML形式データ | 8a8f_validation.xml.gz | 16.1 KB | 表示 | |
| CIF形式データ | 8a8f_validation.cif.gz | 22.5 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/a8/8a8fftp://data.pdbj.org/pub/pdb/validation_reports/a8/8a8f | HTTPS FTP |
-関連構造データ




-リンク
 PDBj
PDBj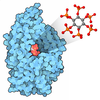
- 集合体
集合体
| 登録構造単位 | 
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||
| 単位格子 |
|
-要素
| #1: タンパク質 | 分子量: 36036.344 Da / 分子数: 1 / 由来タイプ: 組換発現 詳細: The Glc7 C-terminus is covalently linked to chain B (Ref2 348-405) 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: GLC7, CID1, DIS2, YER133W / プラスミド: pET24a 発現宿主:  参照: UniProt: P32598, protein-serine/threonine phosphatase | ||||||
|---|---|---|---|---|---|---|---|
| #2: タンパク質 | 分子量: 8500.543 Da / 分子数: 1 / 由来タイプ: 組換発現 詳細: To trap the Glc7 substrate in the active site, two RNA Pol II CTD repeats were fused C-terminally of Ref2 348-405. The substrate was not visible in the crystal structure. 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: REF2, YDR195W, YD9346.06 発現宿主: 参照: UniProt: P42073 | ||||||
| #3: 化合物 |   分子量: 54.938 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mn 分子量: 54.938 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mn#4: 化合物 | ChemComp-PO4 / | 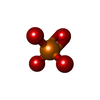  分子量: 94.971 Da / 分子数: 1 / 由来タイプ: 合成 / 式: PO4 分子量: 94.971 Da / 分子数: 1 / 由来タイプ: 合成 / 式: PO4#5: 水 | ChemComp-HOH / |  分子量: 18.015 Da / 分子数: 121 / 由来タイプ: 天然 / 式: H2O 分子量: 18.015 Da / 分子数: 121 / 由来タイプ: 天然 / 式: H2O研究の焦点であるリガンドがあるか | N | |
-実験情報
-実験
| 実験 | 手法: X線回折 / 使用した結晶の数: 1 |
|---|
- 試料調製
試料調製
| 結晶 | マシュー密度: 2.67 Å3/Da / 溶媒含有率: 53.91 % |
|---|---|
| 結晶化 | 温度: 293 K / 手法: 蒸気拡散法, ハンギングドロップ法 / pH: 8 / 詳細: 40 % (v/v) 1,4-Butanediol and 0.1 M Tris-HCl pH 8.5 / PH範囲: 8.5 |
-データ収集
| 回折 | 平均測定温度: 100 K / Serial crystal experiment: N |
|---|---|
| 放射光源 | 由来: シンクロトロン / サイト: Diamond / ビームライン: I04 / 波長: 0.9795 Å |
| 検出器 | タイプ: DECTRIS EIGER2 XE 16M / 検出器: PIXEL / 日付: 2020年12月16日 |
| 放射 | プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray |
| 放射波長 | 波長: 0.9795 Å / 相対比: 1 |
| 反射 | 解像度: 1.85→45.12 Å / Num. obs: 41059 / % possible obs: 99.96 % / 冗長度: 10 % / CC1/2: 1 / Rmerge(I) obs: 0.04671 / Net I/σ(I): 20.46 |
| 反射 シェル | 解像度: 1.85→1.917 Å / 冗長度: 10 % / Rmerge(I) obs: 1.224 / Mean I/σ(I) obs: 1.84 / Num. unique obs: 4028 / CC1/2: 0.9 / % possible all: 98.82 |
- 解析
解析
| ソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 精密化 | 構造決定の手法: 分子置換 開始モデル: 7QWJ 解像度: 1.85→45.12 Å / SU ML: 0.24 / 交差検証法: THROUGHOUT / σ(F): 1.35 / 位相誤差: 28.08 / 立体化学のターゲット値: ML
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | 減衰半径: 0.9 Å / VDWプローブ半径: 1.11 Å / 溶媒モデル: FLAT BULK SOLVENT MODEL | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso max: 122.67 Å2 / Biso mean: 54.6302 Å2 / Biso min: 33.34 Å2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: final / 解像度: 1.85→45.12 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS精密化 シェル | Refine-ID: X-RAY DIFFRACTION / Rfactor Rfree error: 0 / Total num. of bins used: 15
|
