 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-8a8f: Crystal structure of Glc7 phosphatase in complex with the regulat... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8a8f | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Crystal structure of Glc7 phosphatase in complex with the regulatory region of Ref2 | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | GENE REGULATION / Ser/Thr phosphatase / regulatory subunit / RNA / transcription termination | |||||||||
| Function / homology |  Function and homology information Function and homology informationregulation of cell budding / regulation of protein localization to cell division site involved in cytokinesis / cellular bud neck septin collar / mating projection base / regulation of cellular response to glucose starvation / positive regulation of mitotic actomyosin contractile ring assembly / Resolution of Sister Chromatid Cohesion / termination of RNA polymerase II transcription, poly(A)-coupled / chitin biosynthetic process / regulation of glycogen biosynthetic process ...regulation of cell budding / regulation of protein localization to cell division site involved in cytokinesis / cellular bud neck septin collar / mating projection base / regulation of cellular response to glucose starvation / positive regulation of mitotic actomyosin contractile ring assembly / Resolution of Sister Chromatid Cohesion / termination of RNA polymerase II transcription, poly(A)-coupled / chitin biosynthetic process / regulation of glycogen biosynthetic process / cellular bud site selection / positive regulation of clathrin-dependent endocytosis / sno(s)RNA 3'-end processing / positive regulation of exit from mitosis / ascospore formation / protein phosphatase type 1 complex / incipient cellular bud site / negative regulation of actin filament polymerization / intracellular monoatomic ion homeostasis / mRNA cleavage and polyadenylation specificity factor complex / cellular bud neck / protein localization to kinetochore / DNA replication checkpoint signaling / spindle pole body / mRNA 3'-end processing / glycogen metabolic process / cell division site / mitotic spindle assembly checkpoint signaling / protein-serine/threonine phosphatase / protein serine/threonine phosphatase activity / replication fork processing / chromosome organization / response to unfolded protein / DNA damage checkpoint signaling / regulation of mitotic cell cycle / telomere maintenance / chromosome segregation / meiotic cell cycle / kinetochore / mRNA processing / regulation of cell shape / response to heat / regulation of cell cycle / chromatin binding / nucleolus / positive regulation of transcription by RNA polymerase II / RNA binding / metal ion binding / nucleus / cytosol / cytoplasm Similarity search - Function | |||||||||
| Biological species |  | |||||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 1.85 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 1.85 Å | |||||||||
 Authors Authors | Carminati, M. / Manav, C.M. / Bellini, D. / Passmore, L.A. | |||||||||
| Funding support | European Union, 2items
| |||||||||
 Citation Citation | Journal: Mol Cell / Year: 2023 Title: A direct interaction between CPF and RNA Pol II links RNA 3' end processing to transcription. Authors: Manuel Carminati / Juan B Rodríguez-Molina / M Cemre Manav / Dom Bellini / Lori A Passmore /  Abstract: Transcription termination by RNA polymerase II (RNA Pol II) is linked to RNA 3' end processing by the cleavage and polyadenylation factor (CPF or CPSF). CPF contains endonuclease, poly(A) polymerase, ...Transcription termination by RNA polymerase II (RNA Pol II) is linked to RNA 3' end processing by the cleavage and polyadenylation factor (CPF or CPSF). CPF contains endonuclease, poly(A) polymerase, and protein phosphatase activities, which cleave and polyadenylate pre-mRNAs and dephosphorylate RNA Pol II to control transcription. Exactly how the RNA 3' end processing machinery is coupled to transcription remains unclear. Here, we combine in vitro reconstitution, structural studies, and genome-wide analyses to show that yeast CPF physically and functionally interacts with RNA Pol II. Surprisingly, CPF-mediated dephosphorylation promotes the formation of an RNA Pol II stalk-to-stalk homodimer in vitro. This dimer is compatible with transcription but not with the binding of transcription elongation factors. Disruption of the dimerization interface in cells causes transcription defects, including altered RNA Pol II abundance on protein-coding genes, tRNA genes, and intergenic regions. We hypothesize that RNA Pol II dimerization may provide a mechanistic basis for the allosteric model of transcription termination. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8a8f.cif.gz | 90.8 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8a8f.ent.gz | 66.2 KB | Display | PDB format |
| PDBx/mmJSON format | 8a8f.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/a8/8a8fftp://data.pdbj.org/pub/pdb/validation_reports/a8/8a8f | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  7qwjS S: Starting model for refinement C: citing same article ( |
|---|---|
| Similar structure data |



-Links
 PDBj
PDBj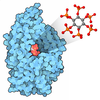
- Assembly
Assembly
| Deposited unit | 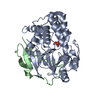
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||
| Unit cell |
|
-Components
| #1: Protein | Mass: 36036.344 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Details: The Glc7 C-terminus is covalently linked to chain B (Ref2 348-405) Source: (gene. exp.) Strain: ATCC 204508 / S288c / Gene: GLC7, CID1, DIS2, YER133W / Plasmid: pET24a Production host:  References: UniProt: P32598, protein-serine/threonine phosphatase | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| #2: Protein | Mass: 8500.543 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Details: To trap the Glc7 substrate in the active site, two RNA Pol II CTD repeats were fused C-terminally of Ref2 348-405. The substrate was not visible in the crystal structure. Source: (gene. exp.) Strain: ATCC 204508 / S288c / Gene: REF2, YDR195W, YD9346.06 Production host: References: UniProt: P42073 | ||||||||
| #3: Chemical |   Mass: 54.938 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Mn Mass: 54.938 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Mn#4: Chemical | ChemComp-PO4 / | 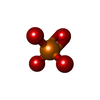  Mass: 94.971 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: PO4 Mass: 94.971 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: PO4#5: Water | ChemComp-HOH / |  Mass: 18.015 Da / Num. of mol.: 121 / Source method: isolated from a natural source / Formula: H2O Mass: 18.015 Da / Num. of mol.: 121 / Source method: isolated from a natural source / Formula: H2OHas ligand of interest | N | Has protein modification | N | |
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 2.67 Å3/Da / Density % sol: 53.91 % |
|---|---|
| Crystal grow | Temperature: 293 K / Method: vapor diffusion, hanging drop / pH: 8 Details: 40 % (v/v) 1,4-Butanediol and 0.1 M Tris-HCl pH 8.5 PH range: 8.5 |
-Data collection
| Diffraction | Mean temperature: 100 K / Serial crystal experiment: N |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: Diamond / Beamline: I04 / Wavelength: 0.9795 Å |
| Detector | Type: DECTRIS EIGER2 XE 16M / Detector: PIXEL / Date: Dec 16, 2020 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 0.9795 Å / Relative weight: 1 |
| Reflection | Resolution: 1.85→45.12 Å / Num. obs: 41059 / % possible obs: 99.96 % / Redundancy: 10 % / CC1/2: 1 / Rmerge(I) obs: 0.04671 / Net I/σ(I): 20.46 |
| Reflection shell | Resolution: 1.85→1.917 Å / Redundancy: 10 % / Rmerge(I) obs: 1.224 / Mean I/σ(I) obs: 1.84 / Num. unique obs: 4028 / CC1/2: 0.9 / % possible all: 98.82 |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: 7QWJ Resolution: 1.85→45.12 Å / SU ML: 0.24 / Cross valid method: THROUGHOUT / σ(F): 1.35 / Phase error: 28.08 / Stereochemistry target values: ML
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.9 Å / VDW probe radii: 1.11 Å / Solvent model: FLAT BULK SOLVENT MODEL | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso max: 122.67 Å2 / Biso mean: 54.6302 Å2 / Biso min: 33.34 Å2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: final / Resolution: 1.85→45.12 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell | Refine-ID: X-RAY DIFFRACTION / Rfactor Rfree error: 0 / Total num. of bins used: 15
|
