 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 7ls1 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | 80S ribosome from mouse bound to eEF2 (Class II) | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード | RIBOSOME / regulation / translation | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報Peptide chain elongation / Synthesis of diphthamide-EEF2 / 5.8S rRNA binding / TNFR1-mediated ceramide production / Protein hydroxylation / positive regulation of amide metabolic process / Formation of the ternary complex, and subsequently, the 43S complex / APC/C:Cdc20 mediated degradation of Cyclin B / SCF-beta-TrCP mediated degradation of Emi1 / APC-Cdc20 mediated degradation of Nek2A ...Peptide chain elongation / Synthesis of diphthamide-EEF2 / 5.8S rRNA binding / TNFR1-mediated ceramide production / Protein hydroxylation / positive regulation of amide metabolic process / Formation of the ternary complex, and subsequently, the 43S complex / APC/C:Cdc20 mediated degradation of Cyclin B / SCF-beta-TrCP mediated degradation of Emi1 / APC-Cdc20 mediated degradation of Nek2A / ER Quality Control Compartment (ERQC) / Regulation of PTEN localization / Regulation of pyruvate metabolism / Downregulation of ERBB2:ERBB3 signaling / IRAK2 mediated activation of TAK1 complex / SMAD2/SMAD3:SMAD4 heterotrimer regulates transcription / PTK6 Regulates RTKs and Their Effectors AKT1 and DOK1 / Gap-filling DNA repair synthesis and ligation in GG-NER / Fanconi Anemia Pathway / Endosomal Sorting Complex Required For Transport (ESCRT) / Negative regulation of FLT3 / Synthesis of active ubiquitin: roles of E1 and E2 enzymes / Regulation of expression of SLITs and ROBOs / IRAK1 recruits IKK complex / IRAK1 recruits IKK complex upon TLR7/8 or 9 stimulation / Downregulation of ERBB4 signaling / E3 ubiquitin ligases ubiquitinate target proteins / Downregulation of TGF-beta receptor signaling / TGF-beta receptor signaling in EMT (epithelial to mesenchymal transition) / Stabilization of p53 / NOTCH3 Activation and Transmission of Signal to the Nucleus / Negative regulators of DDX58/IFIH1 signaling / Alpha-protein kinase 1 signaling pathway / Pexophagy / JNK (c-Jun kinases) phosphorylation and activation mediated by activated human TAK1 / Translesion synthesis by REV1 / Downregulation of SMAD2/3:SMAD4 transcriptional activity / Negative regulation of FGFR3 signaling / Negative regulation of FGFR4 signaling / Translesion synthesis by POLK / Regulation of NF-kappa B signaling / Negative regulation of FGFR1 signaling / Negative regulation of FGFR2 signaling / Regulation of TP53 Activity through Methylation / Formation of a pool of free 40S subunits / NRIF signals cell death from the nucleus / Translesion synthesis by POLI / Regulation of BACH1 activity / Recognition of DNA damage by PCNA-containing replication complex / p75NTR recruits signalling complexes / Interferon alpha/beta signaling / Negative regulation of MAPK pathway / Spry regulation of FGF signaling / SRP-dependent cotranslational protein targeting to membrane / Regulation of TP53 Degradation / Translesion Synthesis by POLH / Activated NOTCH1 Transmits Signal to the Nucleus / Formation of TC-NER Pre-Incision Complex / Major pathway of rRNA processing in the nucleolus and cytosol / Negative regulation of MET activity / TRAF6-mediated induction of TAK1 complex within TLR4 complex / IRAK2 mediated activation of TAK1 complex upon TLR7/8 or 9 stimulation / Termination of translesion DNA synthesis / Nonsense Mediated Decay (NMD) independent of the Exon Junction Complex (EJC) / Autodegradation of Cdh1 by Cdh1:APC/C / APC/C:Cdc20 mediated degradation of Securin / Senescence-Associated Secretory Phenotype (SASP) / Josephin domain DUBs / DNA Damage Recognition in GG-NER / Dual Incision in GG-NER / Ubiquitin-Mediated Degradation of Phosphorylated Cdc25A / Ubiquitin-dependent degradation of Cyclin D / Regulation of TBK1, IKKε (IKBKE)-mediated activation of IRF3, IRF7 / AUF1 (hnRNP D0) binds and destabilizes mRNA / Downregulation of ERBB2 signaling / Dual incision in TC-NER / Oncogene Induced Senescence / PINK1-PRKN Mediated Mitophagy / TNFR1-induced NF-kappa-B signaling pathway / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / Cdc20:Phospho-APC/C mediated degradation of Cyclin A / SCF(Skp2)-mediated degradation of p27/p21 / N-glycan trimming in the ER and Calnexin/Calreticulin cycle / Assembly of the pre-replicative complex / CDK-mediated phosphorylation and removal of Cdc6 / HDR through Homologous Recombination (HRR) / Gap-filling DNA repair synthesis and ligation in TC-NER / Inactivation of CSF3 (G-CSF) signaling / TCF dependent signaling in response to WNT / Metalloprotease DUBs / Formation of Incision Complex in GG-NER / Activation of IRF3, IRF7 mediated by TBK1, IKKε (IKBKE) / EGFR downregulation / Translation initiation complex formation / Ribosomal scanning and start codon recognition / mammalian oogenesis stage / translation at postsynapse / Autodegradation of the E3 ubiquitin ligase COP1 / MAP3K8 (TPL2)-dependent MAPK1/3 activation / G2/M Checkpoints 類似検索 - 分子機能 | |||||||||
| 生物種 |  | |||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.3 Å | |||||||||
 データ登録者 データ登録者 | Loerch, S. / Smith, P.R. / Kunder, N. / Stanowick, A.D. / Lou, T.-F. / Campbell, Z.T. | |||||||||
| 資金援助 |  米国, 2件 米国, 2件
| |||||||||
 引用 引用 | ジャーナル: Nat Commun / 年: 2021 タイトル: Functionally distinct roles for eEF2K in the control of ribosome availability and p-body abundance. 著者: Patrick R Smith / Sarah Loerch / Nikesh Kunder / Alexander D Stanowick / Tzu-Fang Lou / Zachary T Campbell / 要旨: Processing bodies (p-bodies) are a prototypical phase-separated RNA-containing granule. Their abundance is highly dynamic and has been linked to translation. Yet, the molecular mechanisms responsible ...Processing bodies (p-bodies) are a prototypical phase-separated RNA-containing granule. Their abundance is highly dynamic and has been linked to translation. Yet, the molecular mechanisms responsible for coordinate control of the two processes are unclear. Here, we uncover key roles for eEF2 kinase (eEF2K) in the control of ribosome availability and p-body abundance. eEF2K acts on a sole known substrate, eEF2, to inhibit translation. We find that the eEF2K agonist nelfinavir abolishes p-bodies in sensory neurons and impairs translation. To probe the latter, we used cryo-electron microscopy. Nelfinavir stabilizes vacant 80S ribosomes. They contain SERBP1 in place of mRNA and eEF2 in the acceptor site. Phosphorylated eEF2 associates with inactive ribosomes that resist splitting in vitro. Collectively, the data suggest that eEF2K defines a population of inactive ribosomes resistant to recycling and protected from degradation. Thus, eEF2K activity is central to both p-body abundance and ribosome availability in sensory neurons. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7ls1.cif.gz | 5 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7ls1.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 7ls1.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 7ls1_validation.pdf.gz | 1.8 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 7ls1_full_validation.pdf.gz | 2 MB | 表示 | |
| XML形式データ | 7ls1_validation.xml.gz | 350.5 KB | 表示 | |
| CIF形式データ | 7ls1_validation.cif.gz | 616 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/ls/7ls1ftp://data.pdbj.org/pub/pdb/validation_reports/ls/7ls1 | HTTPS FTP |
-関連構造データ







-リンク
 PDBj
PDBj









































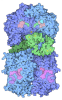








- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
+60S ribosomal protein ... , 43種, 43分子 A1B1C1D1E1F1G1H1D2E2F2G2H2I2J2K2L2M2N2O2P2Q2R2S2T2U2V2W2X2Y2...
-RNA鎖 , 5種, 5分子 A2B2C2m2n2
| #9: RNA鎖 | 分子量: 1532193.125 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #10: RNA鎖 | 分子量: 38998.078 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #11: RNA鎖 | 分子量: 55920.031 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #46: RNA鎖 | 分子量: 603714.625 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #47: RNA鎖 | 分子量: 24231.510 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-タンパク質 , 6種, 6分子 g2I3U3mjA
| #41: タンパク質 | 分子量: 14771.411 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #66: タンパク質 | 分子量: 35115.652 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #78: タンパク質 | 分子量: 17990.014 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #79: タンパク質 | 分子量: 95581.312 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #80: タンパク質 | 分子量: 34252.332 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #82: タンパク質 | 分子量: 42317.527 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
+40S ribosomal protein ... , 31種, 31分子 p2q2r2w2z2o2s2v2x2y2A3B3C3D3E3F3G3H3J3K3L3M3N3O3P3Q3R3S3T3tu
-非ポリマー , 4種, 11分子 

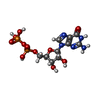

| #86: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 7 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 7 / 由来タイプ: 合成 / 式: Zn#87: 化合物 | ChemComp-MG / |  分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg#88: 化合物 | ChemComp-GDP / |  タイプ: RNA linking / 分子量: 443.201 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H15N5O11P2 / コメント: GDP, エネルギー貯蔵分子*YM タイプ: RNA linking / 分子量: 443.201 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H15N5O11P2 / コメント: GDP, エネルギー貯蔵分子*YM#89: 水 | ChemComp-HOH / | 分子量: 18.015 Da / 分子数: 2 / 由来タイプ: 天然 / 式: H2O |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | N |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: 80S ribosome from mous ebound to eEF2 / タイプ: RIBOSOME / Entity ID: #1-#78, #80-#85 / 由来: NATURAL |
|---|---|
| 分子量 | 値: 4.5 MDa / 実験値: NO |
| 由来(天然) | 生物種: |
| 緩衝液 | pH: 7.5 |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 90 % / 凍結前の試料温度: 277.15 K / 詳細: 3 second blotting, force 3 |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 電子線照射量: 1 e/Å2 / フィルム・検出器のモデル: GATAN K3 (6k x 4k) |
| 電子光学装置 | エネルギーフィルタースリット幅: 20 eV |
- 解析
解析
| ソフトウェア |
| ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 193794 | ||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.3 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 11878 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT / 空間: REAL | ||||||||||||||||||||||||||||||||||||
| 精密化 | 交差検証法: NONE 立体化学のターゲット値: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 160.78 Å2 | ||||||||||||||||||||||||||||||||||||
| 拘束条件 |
|
