 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
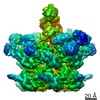
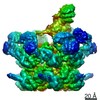
| 登録情報 | データベース: PDB / ID: 7jqq | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | The bacteriophage Phi-29 viral genome packaging motor assembly | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード | MOTOR PROTEIN / packaging motor / ATPase | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報viral DNA genome packaging / 加水分解酵素; 酸無水物に作用; 酸無水物に作用・細胞または細胞小器官の運動に関与 / ATP hydrolysis activity / DNA binding / RNA binding / ATP binding 類似検索 - 分子機能 | |||||||||
| 生物種 |   Bacillus phage phi29 (ファージ)Bacillus virus phi29 (ウイルス) Bacillus phage phi29 (ファージ)Bacillus virus phi29 (ウイルス) | |||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 4.1 Å | |||||||||
 データ登録者 データ登録者 | White, M.A. / Woodson, M. / Morais, M.C. | |||||||||
| 資金援助 |  米国, 2件 米国, 2件
| |||||||||
 引用 引用 | ジャーナル: Sci Adv / 年: 2021 タイトル: A viral genome packaging motor transitions between cyclic and helical symmetry to translocate dsDNA. 著者: Michael Woodson / Joshua Pajak / Bryon P Mahler / Wei Zhao / Wei Zhang / Gaurav Arya / Mark A White / Paul J Jardine / Marc C Morais / 要旨: Molecular segregation and biopolymer manipulation require the action of molecular motors to do work by applying directional forces to macromolecules. The additional strand conserved E (ASCE) ring ...Molecular segregation and biopolymer manipulation require the action of molecular motors to do work by applying directional forces to macromolecules. The additional strand conserved E (ASCE) ring motors are an ancient family of molecular motors responsible for diverse biological polymer manipulation tasks. Viruses use ASCE segregation motors to package their genomes into their protein capsids and provide accessible experimental systems due to their relative simplicity. We show by cryo-EM-focused image reconstruction that ASCE ATPases in viral double-stranded DNA (dsDNA) packaging motors adopt helical symmetry complementary to their dsDNA substrates. Together with previous data, our results suggest that these motors cycle between helical and planar configurations, providing a possible mechanism for directional translocation of DNA. Similar changes in quaternary structure have been observed for proteasome and helicase motors, suggesting an ancient and common mechanism of force generation that has been adapted for specific tasks over the course of evolution. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7jqq.cif.gz | 644.4 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7jqq.ent.gz | 498.1 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 7jqq.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 7jqq_validation.pdf.gz | 1.4 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 7jqq_full_validation.pdf.gz | 1.5 MB | 表示 | |
| XML形式データ | 7jqq_validation.xml.gz | 81.1 KB | 表示 | |
| CIF形式データ | 7jqq_validation.cif.gz | 124.1 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/jq/7jqqftp://data.pdbj.org/pub/pdb/validation_reports/jq/7jqq | HTTPS FTP |
-関連構造データ

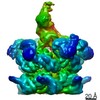
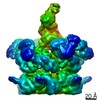
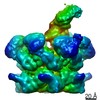

-リンク
 PDBj
PDBj































































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-DNA鎖 , 2種, 2分子 FG
| #3: DNA鎖 | 分子量: 18491.848 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Bacillus virus phi29 (ウイルス) |
|---|---|
| #4: DNA鎖 | 分子量: 18491.850 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Bacillus virus phi29 (ウイルス) / 発現宿主: unidentified (未定義) |
-タンパク質 / RNA鎖 , 2種, 10分子 ABCDEKLMNO
| #1: タンパク質 | 分子量: 39010.406 Da / 分子数: 5 / 由来タイプ: 組換発現 / 由来: (組換発現) Bacillus phage phi29 (ファージ) / 遺伝子: 16 / プラスミド: pSUMO-SacI-XhoI / 発現宿主:  参照: UniProt: P11014, 加水分解酵素; 酸無水物に作用; 酸無水物に作用・細胞または細胞小器官の運動に関与 #2: RNA鎖 | 分子量: 37469.012 Da / 分子数: 5 / 由来タイプ: 組換発現 / 由来: (組換発現) Bacillus virus phi29 (ウイルス) / 発現宿主: unidentified (未定義) |
|---|
-非ポリマー , 2種, 6分子 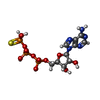

| #5: 化合物 |  分子量: 523.247 Da / 分子数: 3 / 由来タイプ: 合成 / 式: C10H16N5O12P3S 分子量: 523.247 Da / 分子数: 3 / 由来タイプ: 合成 / 式: C10H16N5O12P3Sコメント: ATP-gamma-S, エネルギー貯蔵分子類似体*YM #6: 化合物 |  分子量: 24.305 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Mg |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | N |
|---|---|
| 配列の詳細 | The authors state that the genomic DNA sequences are unknown and the complementary GTCA repeat ...The authors state that the genomic DNA sequences are unknown and the complementary GTCA repeat sequences are just placeholders. |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Bacillus virus phi29 viral genome packaging motor assembly タイプ: VIRUS / Entity ID: #1-#4 / 由来: MULTIPLE SOURCES | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 実験値: NO | ||||||||||||||||||||
| 由来(天然) | 生物種: Bacillus virus phi29 (ウイルス) | ||||||||||||||||||||
| ウイルスについての詳細 | 中空か: NO / エンベロープを持つか: NO / 単離: SPECIES / タイプ: VIRION | ||||||||||||||||||||
| 天然宿主 | 生物種: Bacillus subtilis / 株: A12 | ||||||||||||||||||||
| 緩衝液 | pH: 7.8 | ||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||
| 試料支持 | グリッドの材料: COPPER / グリッドのタイプ: Quantifoil | ||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: TFS KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 最大 デフォーカス(公称値): 4200 nm / 最小 デフォーカス(公称値): 1000 nm / Calibrated defocus min: 1000 nm / 最大 デフォーカス(補正後): 4200 nm / Cs: 2.7 mm / アライメント法: COMA FREE |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 電子線照射量: 41.4 e/Å2 / 検出モード: SUPER-RESOLUTION フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) |
| 電子光学装置 | エネルギーフィルター名称: GIF 2002 |
| 画像スキャン | 横: 7676 / 縦: 7420 / 動画フレーム数/画像: 45 / 利用したフレーム数/画像: 0-44 |
- 解析
解析
| ソフトウェア |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING ONLY | ||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||
| 3次元再構成 | 解像度: 4.1 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 12526 / アルゴリズム: BACK PROJECTION 詳細: The final step before reconstruction was classification without alignment, which was done with even/odd sets mixed together, but even/odd were completely independent for all angular assignment steps クラス平均像の数: 1 / 対称性のタイプ: POINT | ||||||||||||||||||||||||
| 精密化 | 交差検証法: NONE 立体化学のターゲット値: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 55.8 Å2 | ||||||||||||||||||||||||
| 拘束条件 |
|
