 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 7a08 | |||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | CryoEM Structure of cGAS Nucleosome complex | |||||||||||||||||||||||||||||||||||||||||||||
 Components Components |
| |||||||||||||||||||||||||||||||||||||||||||||
 Keywords Keywords | TRANSFERASE / complex / immune protein / nuclear protein | |||||||||||||||||||||||||||||||||||||||||||||
| Function / homology |  Function and homology information Function and homology information: / Barr body / 2',3'-cyclic GMP-AMP synthase activity / cyclic GMP-AMP synthase / regulation of type I interferon production / negative regulation of chromosome condensation / paracrine signaling / pericentric heterochromatin formation / inner kinetochore / poly-ADP-D-ribose modification-dependent protein binding ...: / Barr body / 2',3'-cyclic GMP-AMP synthase activity / cyclic GMP-AMP synthase / regulation of type I interferon production / negative regulation of chromosome condensation / paracrine signaling / pericentric heterochromatin formation / inner kinetochore / poly-ADP-D-ribose modification-dependent protein binding / regulation of immunoglobulin production / muscle cell differentiation / : / oocyte maturation / regulation of T cell activation / cGAS/STING signaling pathway / nucleosomal DNA binding / pattern recognition receptor signaling pathway / negative regulation of DNA repair / negative regulation of cGAS/STING signaling pathway / nucleus organization / cellular response to exogenous dsRNA / cytoplasmic pattern recognition receptor signaling pathway / spermatid development / regulation of immune response / single fertilization / subtelomeric heterochromatin formation / negative regulation of double-strand break repair via homologous recombination / RNA polymerase II core promoter sequence-specific DNA binding / nucleosome binding / negative regulation of megakaryocyte differentiation / protein localization to CENP-A containing chromatin / positive regulation of defense response to virus by host / positive regulation of type I interferon production / Replacement of protamines by nucleosomes in the male pronucleus / CENP-A containing nucleosome / phosphatidylinositol-4,5-bisphosphate binding / Packaging Of Telomere Ends / embryo implantation / activation of innate immune response / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / Deposition of new CENPA-containing nucleosomes at the centromere / telomere organization / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / RNA Polymerase I Promoter Opening / Inhibition of DNA recombination at telomere / Assembly of the ORC complex at the origin of replication / Meiotic synapsis / SUMOylation of chromatin organization proteins / Regulation of endogenous retroelements by the Human Silencing Hub (HUSH) complex / DNA methylation / Condensation of Prophase Chromosomes / determination of adult lifespan / Chromatin modifications during the maternal to zygotic transition (MZT) / SIRT1 negatively regulates rRNA expression / HCMV Late Events / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / PRC2 methylates histones and DNA / Regulation of endogenous retroelements by KRAB-ZFP proteins / Defective pyroptosis / innate immune response in mucosa / HDACs deacetylate histones / Regulation of endogenous retroelements by Piwi-interacting RNAs (piRNAs) / molecular condensate scaffold activity / RNA Polymerase I Promoter Escape / Nonhomologous End-Joining (NHEJ) / Transcriptional regulation by small RNAs / HDMs demethylate histones / Formation of the beta-catenin:TCF transactivating complex / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / NoRC negatively regulates rRNA expression / Negative Regulation of CDH1 Gene Transcription / G2/M DNA damage checkpoint / PKMTs methylate histone lysines / B-WICH complex positively regulates rRNA expression / DNA Damage/Telomere Stress Induced Senescence / male gonad development / Meiotic recombination / Pre-NOTCH Transcription and Translation / multicellular organism growth / Activation of anterior HOX genes in hindbrain development during early embryogenesis / Transcriptional regulation of granulopoiesis / RMTs methylate histone arginines / Metalloprotease DUBs / HCMV Early Events / osteoblast differentiation / structural constituent of chromatin / positive regulation of cellular senescence / UCH proteinases / nucleosome / antimicrobial humoral immune response mediated by antimicrobial peptide / nucleosome assembly / heterochromatin formation / HATs acetylate histones / antibacterial humoral response / E3 ubiquitin ligases ubiquitinate target proteins / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks Similarity search - Function | |||||||||||||||||||||||||||||||||||||||||||||
| Biological species |   Homo sapiens (human) Homo sapiens (human)synthetic construct (others) | |||||||||||||||||||||||||||||||||||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.11 Å | |||||||||||||||||||||||||||||||||||||||||||||
 Authors Authors | Michalski, S. / de Oliveira Mann, C.C. / Witte, G. / Bartho, J. / Lammens, K. / Hopfner, K.P. | |||||||||||||||||||||||||||||||||||||||||||||
| Funding support |  Germany, 1items Germany, 1items
| |||||||||||||||||||||||||||||||||||||||||||||
 Citation Citation | Journal: Nature / Year: 2020 Title: Structural basis for sequestration and autoinhibition of cGAS by chromatin. Authors: Sebastian Michalski / Carina C de Oliveira Mann / Che A Stafford / Gregor Witte / Joseph Bartho / Katja Lammens / Veit Hornung / Karl-Peter Hopfner / Abstract: Cyclic GMP-AMP synthase (cGAS) is an innate immune sensor for cytosolic microbial DNA. After binding DNA, cGAS synthesizes the messenger 2'3'-cyclic GMP-AMP (cGAMP), which triggers cell-autonomous ...Cyclic GMP-AMP synthase (cGAS) is an innate immune sensor for cytosolic microbial DNA. After binding DNA, cGAS synthesizes the messenger 2'3'-cyclic GMP-AMP (cGAMP), which triggers cell-autonomous defence and the production of type I interferons and pro-inflammatory cytokines via the activation of STING. In addition to responding to cytosolic microbial DNA, cGAS also recognizes mislocalized cytosolic self-DNA and has been implicated in autoimmunity and sterile inflammation. Specificity towards pathogen- or damage-associated DNA was thought to be caused by cytosolic confinement. However, recent findings place cGAS robustly in the nucleus, where tight tethering of chromatin is important to prevent autoreactivity to self-DNA. Here we show how cGAS is sequestered and inhibited by chromatin. We provide a cryo-electron microscopy structure of the cGAS catalytic domain bound to a nucleosome, which shows that cGAS does not interact with the nucleosomal DNA, but instead interacts with histone 2A-histone 2B, and is tightly anchored to the 'acidic patch'. The interaction buries the cGAS DNA-binding site B, and blocks the formation of active cGAS dimers. The acidic patch robustly outcompetes agonistic DNA for binding to cGAS, which suggests that nucleosome sequestration can efficiently inhibit cGAS, even when accessible DNA is nearby, such as in actively transcribed genomic regions. Our results show how nuclear cGAS is sequestered by chromatin and provides a mechanism for preventing autoreactivity to nuclear self-DNA. | |||||||||||||||||||||||||||||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 7a08.cif.gz | 331.7 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb7a08.ent.gz | 248.9 KB | Display | PDB format |
| PDBx/mmJSON format | 7a08.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/a0/7a08ftp://data.pdbj.org/pub/pdb/validation_reports/a0/7a08 | HTTPS FTP |
|---|
-Related structure data
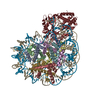
| Related structure data | 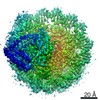 11601MC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |
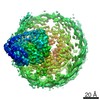
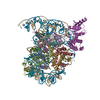
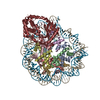
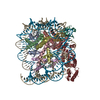
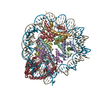
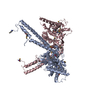
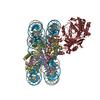
-Links
 PDBj
PDBj










































- Assembly
Assembly
| Deposited unit | 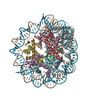
|
|---|---|
| 1 |
|
-Components
-Protein , 5 types, 9 molecules abfcgdhei
| #1: Protein | Mass: 43401.180 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.)  | ||||||
|---|---|---|---|---|---|---|---|
| #4: Protein | Mass: 14004.329 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: H2AC6, H2AFL, HIST1H2AC / Production host: #5: Protein | Mass: 13806.018 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human)Gene: HIST1H2BC, H2BFL, HIST1H2BE, H2BFH, HIST1H2BF, H2BFG, HIST1H2BG, H2BFA, HIST1H2BI, H2BFK Production host: #6: Protein | Mass: 15229.787 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: H3-3A, H3.3A, H3F3, H3F3A, PP781, H3-3B, H3.3B, H3F3B / Production host: #7: Protein | Mass: 11263.231 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human)Gene: HIST1H4A, H4/A, H4FA, HIST1H4B, H4/I, H4FI, HIST1H4C, H4/G, H4FG, HIST1H4D, H4/B, H4FB, HIST1H4E, H4/J, H4FJ, HIST1H4F, H4/C, H4FC, HIST1H4H, H4/H, H4FH, HIST1H4I, H4/M, H4FM, HIST1H4J, H4/E, ...Gene: HIST1H4A, H4/A, H4FA, HIST1H4B, H4/I, H4FI, HIST1H4C, H4/G, H4FG, HIST1H4D, H4/B, H4FB, HIST1H4E, H4/J, H4FJ, HIST1H4F, H4/C, H4FC, HIST1H4H, H4/H, H4FH, HIST1H4I, H4/M, H4FM, HIST1H4J, H4/E, H4FE, HIST1H4K, H4/D, H4FD, HIST1H4L, H4/K, H4FK, HIST2H4A, H4/N, H4F2, H4FN, HIST2H4, HIST2H4B, H4/O, H4FO, HIST4H4 Production host: |
-Nucleosomal DNA strand ... , 2 types, 2 molecules IJ
| #2: DNA chain | Mass: 45145.754 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) synthetic construct (others) |
|---|---|
| #3: DNA chain | Mass: 45604.047 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) synthetic construct (others) |
-Non-polymers , 1 types, 1 molecules 
| #8: Chemical | ChemComp-ZN /  Mass: 65.409 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Zn Mass: 65.409 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Zn |
|---|
-Details
| Has ligand of interest | N |
|---|---|
| Has protein modification | Y |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component |
| ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight |
| ||||||||||||||||||||||||||||||
| Source (natural) |
| ||||||||||||||||||||||||||||||
| Source (recombinant) |
| ||||||||||||||||||||||||||||||
| Buffer solution | pH: 7.5 | ||||||||||||||||||||||||||||||
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||||||||||||
| Specimen support | Grid material: COPPER / Grid type: Quantifoil | ||||||||||||||||||||||||||||||
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD |
| Image recording | Electron dose: 44.8 e/Å2 / Film or detector model: GATAN K2 SUMMIT (4k x 4k) |
- Processing
Processing
| Software | Name: PHENIX / Version: 1.18.2_3874: / Classification: refinement | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM software | Name: PHENIX / Category: model refinement | ||||||||||||||||||||||||
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| 3D reconstruction | Resolution: 3.11 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 172977 / Symmetry type: POINT | ||||||||||||||||||||||||
| Refine LS restraints |
|
