 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 6tny | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
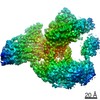
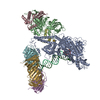
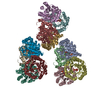
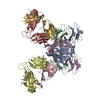
| Title | Processive human polymerase delta holoenzyme | ||||||||||||||||||||||||||||||||||||
 Components Components |
| ||||||||||||||||||||||||||||||||||||
 Keywords Keywords | REPLICATION / Protein | ||||||||||||||||||||||||||||||||||||
| Function / homology |  Function and homology information Function and homology informationdelta DNA polymerase complex / DNA synthesis involved in UV-damage excision repair / zeta DNA polymerase complex / nucleotide-excision repair complex / Cytosolic iron-sulfur cluster assembly / dinucleotide insertion or deletion binding / PCNA-p21 complex / mitotic telomere maintenance via semi-conservative replication / purine-specific mismatch base pair DNA N-glycosylase activity / nucleotide-excision repair, DNA gap filling ...delta DNA polymerase complex / DNA synthesis involved in UV-damage excision repair / zeta DNA polymerase complex / nucleotide-excision repair complex / Cytosolic iron-sulfur cluster assembly / dinucleotide insertion or deletion binding / PCNA-p21 complex / mitotic telomere maintenance via semi-conservative replication / purine-specific mismatch base pair DNA N-glycosylase activity / nucleotide-excision repair, DNA gap filling / nuclear lamina / Polymerase switching / Processive synthesis on the lagging strand / DNA replication proofreading / PCNA complex / MutLalpha complex binding / 3'-5'-DNA exonuclease activity / Removal of the Flap Intermediate / Telomere C-strand (Lagging Strand) Synthesis / Mismatch repair (MMR) directed by MSH2:MSH3 (MutSbeta) / Mismatch repair (MMR) directed by MSH2:MSH6 (MutSalpha) / Transcription of E2F targets under negative control by DREAM complex / Polymerase switching on the C-strand of the telomere / replisome / Processive synthesis on the C-strand of the telomere / response to L-glutamate / aggresome / Removal of the Flap Intermediate from the C-strand / Hydrolases; Acting on ester bonds; Exodeoxyribonucleases producing 5'-phosphomonoesters / error-free translesion synthesis / response to dexamethasone / DNA biosynthetic process / DNA strand elongation involved in DNA replication / histone acetyltransferase binding / leading strand elongation / DNA polymerase processivity factor activity / G1/S-Specific Transcription / DNA synthesis involved in DNA repair / nuclear replication fork / SUMOylation of DNA replication proteins / replication fork processing / PCNA-Dependent Long Patch Base Excision Repair / error-prone translesion synthesis / fatty acid homeostasis / response to cadmium ion / estrous cycle / response to UV / mismatch repair / cyclin-dependent protein kinase holoenzyme complex / base-excision repair, gap-filling / translesion synthesis / epithelial cell differentiation / DNA polymerase binding / positive regulation of endothelial cell proliferation / TP53 Regulates Transcription of Genes Involved in G2 Cell Cycle Arrest / liver regeneration / positive regulation of DNA replication / nuclear estrogen receptor binding / replication fork / positive regulation of DNA repair / Translesion synthesis by REV1 / Translesion synthesis by POLK / male germ cell nucleus / Translesion synthesis by POLI / Gap-filling DNA repair synthesis and ligation in GG-NER / Termination of translesion DNA synthesis / Translesion Synthesis by POLH / Recognition of DNA damage by PCNA-containing replication complex / receptor tyrosine kinase binding / DNA-templated DNA replication / cellular response to xenobiotic stimulus / HDR through Homologous Recombination (HRR) / Dual Incision in GG-NER / cellular response to hydrogen peroxide / Dual incision in TC-NER / Gap-filling DNA repair synthesis and ligation in TC-NER / cellular response to UV / response to estradiol / E3 ubiquitin ligases ubiquitinate target proteins / heart development / chromatin organization / 4 iron, 4 sulfur cluster binding / DNA-directed DNA polymerase / damaged DNA binding / DNA-directed DNA polymerase activity / protein-macromolecule adaptor activity / DNA replication / chromosome, telomeric region / nuclear body / nucleotide binding / DNA repair / chromatin binding / centrosome / chromatin / protein-containing complex binding / enzyme binding / negative regulation of transcription by RNA polymerase II / DNA binding / extracellular exosome / zinc ion binding Similarity search - Function | ||||||||||||||||||||||||||||||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human)synthetic construct (others) | ||||||||||||||||||||||||||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.08 Å | ||||||||||||||||||||||||||||||||||||
 Authors Authors | Lancey, C. / Hamdan, S.M. / De Biasio, A. | ||||||||||||||||||||||||||||||||||||
| Funding support |  United Kingdom, 1items United Kingdom, 1items
| ||||||||||||||||||||||||||||||||||||
 Citation Citation | Journal: Nat Commun / Year: 2020 Title: Structure of the processive human Pol δ holoenzyme. Authors: Claudia Lancey / Muhammad Tehseen / Vlad-Stefan Raducanu / Fahad Rashid / Nekane Merino / Timothy J Ragan / Christos G Savva / Manal S Zaher / Afnan Shirbini / Francisco J Blanco / Samir M ...Authors: Claudia Lancey / Muhammad Tehseen / Vlad-Stefan Raducanu / Fahad Rashid / Nekane Merino / Timothy J Ragan / Christos G Savva / Manal S Zaher / Afnan Shirbini / Francisco J Blanco / Samir M Hamdan / Alfredo De Biasio /   Abstract: In eukaryotes, DNA polymerase δ (Pol δ) bound to the proliferating cell nuclear antigen (PCNA) replicates the lagging strand and cooperates with flap endonuclease 1 (FEN1) to process the Okazaki ...In eukaryotes, DNA polymerase δ (Pol δ) bound to the proliferating cell nuclear antigen (PCNA) replicates the lagging strand and cooperates with flap endonuclease 1 (FEN1) to process the Okazaki fragments for their ligation. We present the high-resolution cryo-EM structure of the human processive Pol δ-DNA-PCNA complex in the absence and presence of FEN1. Pol δ is anchored to one of the three PCNA monomers through the C-terminal domain of the catalytic subunit. The catalytic core sits on top of PCNA in an open configuration while the regulatory subunits project laterally. This arrangement allows PCNA to thread and stabilize the DNA exiting the catalytic cleft and recruit FEN1 to one unoccupied monomer in a toolbelt fashion. Alternative holoenzyme conformations reveal important functional interactions that maintain PCNA orientation during synthesis. This work sheds light on the structural basis of Pol δ's activity in replicating the human genome. | ||||||||||||||||||||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 6tny.cif.gz | 465.9 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb6tny.ent.gz | 361.5 KB | Display | PDB format |
| PDBx/mmJSON format | 6tny.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/tn/6tnyftp://data.pdbj.org/pub/pdb/validation_reports/tn/6tny | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  10539MC  6s1mC  6s1nC  6s1oC  6tnzC C: citing same article ( M: map data used to model this data |
|---|---|
| Similar structure data | |
| EM raw data | EMPIAR-10823 (Title: Cryo electron micrographs of Pol delta-PCNA-DNA-FEN1 sample Data size: 1.6 TB Data #1: Cryo-electron micrographs of Pol delta-FEN1 toolbelt [micrographs - multiframe]) |






-Links
 PDBj
PDBj







































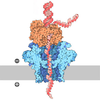







- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-Protein , 2 types, 4 molecules AEFG
| #1: Protein | Mass: 123785.922 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: POLD1, POLD / Cell line (production host): Sf9 / Production host:   Spodoptera frugiperda (fall armyworm) Spodoptera frugiperda (fall armyworm)References: UniProt: P28340, DNA-directed DNA polymerase, Hydrolases; Acting on ester bonds; Exodeoxyribonucleases producing 5'-phosphomonoesters |
|---|---|
| #5: Protein | Mass: 29088.061 Da / Num. of mol.: 3 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: PCNA / Production host:  |
-DNA polymerase delta subunit ... , 3 types, 3 molecules BCD
| #2: Protein | Mass: 51338.168 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: POLD2 / Cell line (production host): Sf9 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: P49005 |
|---|---|
| #3: Protein | Mass: 52528.500 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: POLD3, KIAA0039 / Cell line (production host): Sf9 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: Q15054 |
| #4: Protein | Mass: 16274.323 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: POLD4, POLDS / Cell line (production host): Sf9 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: Q9HCU8 |
-DNA chain , 2 types, 2 molecules PT
| #6: DNA chain | Mass: 7665.987 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) synthetic construct (others) |
|---|---|
| #7: DNA chain | Mass: 11677.539 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) synthetic construct (others) |
-Non-polymers , 3 types, 3 molecules 
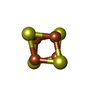

| #8: Chemical | ChemComp-ZN /  Mass: 65.409 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Zn Mass: 65.409 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Zn |
|---|---|
| #9: Chemical | ChemComp-SF4 /  Mass: 351.640 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Fe4S4 Mass: 351.640 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Fe4S4 |
| #10: Chemical | ChemComp-TTP /  Mass: 482.168 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H17N2O14P3 Mass: 482.168 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H17N2O14P3 |
-Details
| Has ligand of interest | N |
|---|---|
| Has protein modification | N |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component |
| |||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Experimental value: NO | |||||||||||||||||||||||||||||||||||
| Source (natural) |
| |||||||||||||||||||||||||||||||||||
| Source (recombinant) |
| |||||||||||||||||||||||||||||||||||
| Buffer solution | pH: 7.5 | |||||||||||||||||||||||||||||||||||
| Buffer component |
| |||||||||||||||||||||||||||||||||||
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES / Details: Complex separated by gel filtration | |||||||||||||||||||||||||||||||||||
| Specimen support | Grid material: GOLD / Grid mesh size: 300 divisions/in. / Grid type: Quantifoil, UltrAuFoil, R1.2/1.3 | |||||||||||||||||||||||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK IV / Cryogen name: ETHANE / Humidity: 100 % / Chamber temperature: 277 K |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 105000 X / Calibrated magnification: 57471 X / Nominal defocus max: 2300 nm / Nominal defocus min: 1100 nm / Cs: 2.7 mm / C2 aperture diameter: 50 µm / Alignment procedure: ZEMLIN TABLEAU |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER / Temperature (max): 77 K / Temperature (min): 77 K |
| Image recording | Average exposure time: 3 sec. / Electron dose: 44 e/Å2 / Film or detector model: GATAN K3 BIOQUANTUM (6k x 4k) / Num. of grids imaged: 1 / Num. of real images: 5071 / Details: Data were collected in super resolution mode |
| Image scans | Width: 11520 / Height: 8184 |
- Processing
Processing
| Software | Name: PHENIX / Version: 1.14_3260: / Classification: refinement | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM software |
| ||||||||||||||||||||
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | ||||||||||||||||||||
| 3D reconstruction | Resolution: 3.08 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 288920 / Symmetry type: POINT |
