 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 6r6h | ||||||
|---|---|---|---|---|---|---|---|
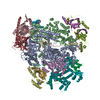
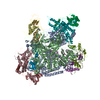
| タイトル | Structural basis of Cullin-2 RING E3 ligase regulation by the COP9 signalosome | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | LIGASE / Cullin-Ring E3 Ligase COP9 Signalosome Neddylation | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報nucleotide-excision repair factor 4 complex / COP9 signalosome assembly / trophectodermal cell proliferation / macrophage migration inhibitory factor binding / regulation of IRE1-mediated unfolded protein response / global genome nucleotide-excision repair / exosomal secretion / GTPase inhibitor activity / deNEDDylase activity / protein deneddylation ...nucleotide-excision repair factor 4 complex / COP9 signalosome assembly / trophectodermal cell proliferation / macrophage migration inhibitory factor binding / regulation of IRE1-mediated unfolded protein response / global genome nucleotide-excision repair / exosomal secretion / GTPase inhibitor activity / deNEDDylase activity / protein deneddylation / eukaryotic translation initiation factor 3 complex / regulation of protein neddylation / activation of NF-kappaB-inducing kinase activity / regulation of cellular response to hypoxia / cullin-RING-type E3 NEDD8 transferase / NEDD8 transferase activity / COP9 signalosome / cullin-RING ubiquitin ligase complex / RHOBTB3 ATPase cycle / negative regulation of receptor signaling pathway via JAK-STAT / Cul7-RING ubiquitin ligase complex / ubiquitin-dependent protein catabolic process via the C-end degron rule pathway / cellular response to chemical stress / Loss of Function of FBXW7 in Cancer and NOTCH1 Signaling / transcription elongation factor activity / target-directed miRNA degradation / elongin complex / positive regulation of protein autoubiquitination / RNA polymerase II transcription initiation surveillance / protein neddylation / metal-dependent deubiquitinase activity / Replication of the SARS-CoV-1 genome / regulation of JNK cascade / NEDD8 ligase activity / 加水分解酵素; プロテアーゼ; ペプチド結合加水分解酵素 / RHOBTB1 GTPase cycle / VCB complex / negative regulation of response to oxidative stress / regulation of DNA damage response, signal transduction by p53 class mediator / Cul5-RING ubiquitin ligase complex / inner cell mass cell proliferation / SCF ubiquitin ligase complex / negative regulation of type I interferon production / ubiquitin-ubiquitin ligase activity / Cul2-RING ubiquitin ligase complex / intracellular membraneless organelle / SCF-dependent proteasomal ubiquitin-dependent protein catabolic process / Cul4A-RING E3 ubiquitin ligase complex / Cul4-RING E3 ubiquitin ligase complex / TP53 Regulates Transcription of DNA Repair Genes / Cul3-RING ubiquitin ligase complex / Cul4B-RING E3 ubiquitin ligase complex / ubiquitin ligase complex scaffold activity / negative regulation of mitophagy / SUMOylation of ubiquitinylation proteins / Prolactin receptor signaling / response to light stimulus / cullin family protein binding / Antigen processing: Ubiquitination & Proteasome degradation / skeletal muscle cell differentiation / Pausing and recovery of Tat-mediated HIV elongation / Tat-mediated HIV elongation arrest and recovery / negative regulation of transcription elongation by RNA polymerase II / HIV elongation arrest and recovery / Pausing and recovery of HIV elongation / protein monoubiquitination / negative regulation of signal transduction / Tat-mediated elongation of the HIV-1 transcript / positive regulation of DNA-binding transcription factor activity / Formation of HIV-1 elongation complex containing HIV-1 Tat / ubiquitin-like ligase-substrate adaptor activity / protein K48-linked ubiquitination / response to UV / Formation of HIV elongation complex in the absence of HIV Tat / Nuclear events stimulated by ALK signaling in cancer / RNA Polymerase II Transcription Elongation / Formation of RNA Pol II elongation complex / JNK cascade / negative regulation of TORC1 signaling / transcription-coupled nucleotide-excision repair / translation initiation factor activity / RNA Polymerase II Pre-transcription Events / positive regulation of TORC1 signaling / regulation of cellular response to insulin stimulus / intrinsic apoptotic signaling pathway / negative regulation of insulin receptor signaling pathway / post-translational protein modification / negative regulation of autophagy / protein serine/threonine kinase binding / Regulation of BACH1 activity / T cell activation / transcription corepressor binding / nucleotide-excision repair / positive regulation of cell differentiation / Degradation of DVL / TP53 Regulates Transcription of DNA Repair Genes / cellular response to amino acid stimulus / transcription initiation at RNA polymerase II promoter / transcription elongation by RNA polymerase II / Degradation of GLI1 by the proteasome 類似検索 - 分子機能 | ||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||
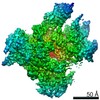
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 8.4 Å | ||||||
 データ登録者 データ登録者 | Morris, E.P. / Faull, S.V. / Lau, A.M.C. / Politis, A. / Beuron, F. / Cronin, N. | ||||||
| 資金援助 |  英国, 1件 英国, 1件
| ||||||
 引用 引用 | ジャーナル: Nat Commun / 年: 2019 タイトル: Structural basis of Cullin 2 RING E3 ligase regulation by the COP9 signalosome. 著者: Sarah V Faull / Andy M C Lau / Chloe Martens / Zainab Ahdash / Kjetil Hansen / Hugo Yebenes / Carla Schmidt / Fabienne Beuron / Nora B Cronin / Edward P Morris / Argyris Politis /   要旨: Cullin-Ring E3 Ligases (CRLs) regulate a multitude of cellular pathways through specific substrate receptors. The COP9 signalosome (CSN) deactivates CRLs by removing NEDD8 from activated Cullins. ...Cullin-Ring E3 Ligases (CRLs) regulate a multitude of cellular pathways through specific substrate receptors. The COP9 signalosome (CSN) deactivates CRLs by removing NEDD8 from activated Cullins. Here we present structures of the neddylated and deneddylated CSN-CRL2 complexes by combining single-particle cryo-electron microscopy (cryo-EM) with chemical cross-linking mass spectrometry (XL-MS). These structures suggest a conserved mechanism of CSN activation, consisting of conformational clamping of the CRL2 substrate by CSN2/CSN4, release of the catalytic CSN5/CSN6 heterodimer and finally activation of the CSN5 deneddylation machinery. Using hydrogen-deuterium exchange (HDX)-MS we show that CRL2 activates CSN5/CSN6 in a neddylation-independent manner. The presence of NEDD8 is required to activate the CSN5 active site. Overall, by synergising cryo-EM with MS, we identify sensory regions of the CSN that mediate its stepwise activation and provide a framework for understanding the regulatory mechanism of other Cullin family members. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 6r6h.cif.gz | 815.7 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb6r6h.ent.gz | 641.5 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 6r6h.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 6r6h_validation.pdf.gz | 893.8 KB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 6r6h_full_validation.pdf.gz | 1.6 MB | 表示 | |
| XML形式データ | 6r6h_validation.xml.gz | 221.1 KB | 表示 | |
| CIF形式データ | 6r6h_validation.cif.gz | 309.3 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/r6/6r6hftp://data.pdbj.org/pub/pdb/validation_reports/r6/6r6h | HTTPS FTP |
-関連構造データ
| 関連構造データ |  4736MC  4739C  4741C  4742C  4744C  6r7fC  6r7hC  6r7iC  6r7nC M: このデータのモデリングに利用したマップデータ C: 同じ文献を引用 ( |
|---|---|
| 類似構造データ |



-リンク
 PDBj
PDBj






















- 集合体
集合体
| 登録構造単位 | 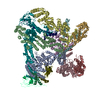
|
|---|---|
| 1 |
|
-要素
-COP9 signalosome complex subunit ... , 8種, 8分子 ABCDEFHG
| #1: タンパク質 | 分子量: 55606.496 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: GPS1, COPS1, CSN1 / 細胞株 (発現宿主): SF9発現宿主:   Spodoptera frugiperda (ツマジロクサヨトウ) Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q13098 |
|---|---|
| #2: タンパク質 | 分子量: 51664.570 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: COPS2, CSN2, TRIP15 / 細胞株 (発現宿主): SF9発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: P61201 |
| #3: タンパク質 | 分子量: 45808.816 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: COPS3, CSN3 / 細胞株 (発現宿主): SF9発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q9UNS2 |
| #4: タンパク質 | 分子量: 46322.688 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: COPS4, CSN4 / 細胞株 (発現宿主): SF9発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q9BT78 |
| #5: タンパク質 | 分子量: 37621.742 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: COPS5, CSN5, JAB1 / 細胞株 (発現宿主): SF9発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q92905, 加水分解酵素; プロテアーゼ; ペプチド結合加水分解酵素 |
| #6: タンパク質 | 分子量: 34713.844 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: COPS6, CSN6, HVIP / 細胞株 (発現宿主): SF9発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q7L5N1 |
| #7: タンパク質 | 分子量: 23245.543 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: COPS8, CSN8 / 細胞株 (発現宿主): SF9発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q99627 |
| #13: タンパク質 | 分子量: 23228.529 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: COPS7B, CSN7B / 細胞株 (発現宿主): SF9発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q9H9Q2 |
-タンパク質 , 5種, 5分子 OPQRV
| #8: タンパク質 | 分子量: 87098.930 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: CUL2 / 細胞株 (発現宿主): SF9発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q13617 |
|---|---|
| #9: タンパク質 | 分子量: 11819.483 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: ELOB, TCEB2 / 細胞株 (発現宿主): SF9発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q15370 |
| #10: タンパク質 | 分子量: 11338.760 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 細胞株 (発現宿主): SF9発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q03071*PLUS |
| #11: タンパク質 | 分子量: 10092.631 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 細胞株 (発現宿主): SF9発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: P62877*PLUS |
| #12: タンパク質 | 分子量: 17330.764 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: VHL / 細胞株 (発現宿主): SF9発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: P40337 |
-非ポリマー , 1種, 1分子 
| #14: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Zn |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Cullin-Ring E3 Ligases (CRLs) complexes with neddylated COP9 signalosome (CSN) タイプ: COMPLEX / Entity ID: #1-#13 / 由来: RECOMBINANT |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 由来(組換発現) | 生物種: Spodoptera frugiperda (ツマジロクサヨトウ) 細胞: sf9 |
| 緩衝液 | pH: 7.5 詳細: 15 mM Hepes pH 7.5 100 mM NaCl 0.5 mM DTT 1% glycerol |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: NITROGEN |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 47170 X / 最大 デフォーカス(公称値): 3000 nm / 最小 デフォーカス(公称値): 1800 nm / Cs: 2.7 mm / C2レンズ絞り径: 100 µm |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 電子線照射量: 45 e/Å2 / 検出モード: COUNTING フィルム・検出器のモデル: GATAN K2 QUANTUM (4k x 4k) |
| 電子光学装置 | エネルギーフィルター名称: GIF Bioquantum / エネルギーフィルタースリット幅: 30 eV |
- 解析
解析
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION |
|---|---|
| 対称性 | 点対称性: C1 (非対称) |
| 3次元再構成 | 解像度: 8.4 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 24040 / 対称性のタイプ: POINT |
