 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-6gct: cryo-EM structure of the human neutral amino acid transporter ASCT2 -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 6gct | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | cryo-EM structure of the human neutral amino acid transporter ASCT2 | ||||||||||||||||||
 Components Components | Neutral amino acid transporter B(0) | ||||||||||||||||||
 Keywords Keywords | MEMBRANE PROTEIN / neutral amino acid transporter | ||||||||||||||||||
| Function / homology |  Function and homology information Function and homology informationL-glutamine secretion / L-glutamine import across plasma membrane / L-glutamine transmembrane transporter activity / L-serine transmembrane transporter activity / L-glutamine transport / ligand-gated channel activity / L-aspartate transmembrane transporter activity / neutral amino acid transport / L-aspartate import across plasma membrane / neutral L-amino acid transmembrane transporter activity ...L-glutamine secretion / L-glutamine import across plasma membrane / L-glutamine transmembrane transporter activity / L-serine transmembrane transporter activity / L-glutamine transport / ligand-gated channel activity / L-aspartate transmembrane transporter activity / neutral amino acid transport / L-aspartate import across plasma membrane / neutral L-amino acid transmembrane transporter activity / symporter activity / Amino acid transport across the plasma membrane / amino acid transmembrane transporter activity / antiporter activity / RHOJ GTPase cycle / protein homotrimerization / RHOQ GTPase cycle / amino acid transport / RHOH GTPase cycle / RAC3 GTPase cycle / transport across blood-brain barrier / centriolar satellite / RAC1 GTPase cycle / basal plasma membrane / erythrocyte differentiation / melanosome / virus receptor activity / signaling receptor activity / ciliary basal body / extracellular exosome / membrane / metal ion binding / plasma membrane Similarity search - Function | ||||||||||||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.85 Å | ||||||||||||||||||
 Authors Authors | Garaeva, A.A. / Oostergetel, G.T. / Gati, C. / Guskov, A. / Paulino, C. / Slotboom, D.J. | ||||||||||||||||||
| Funding support |  Netherlands, 5items Netherlands, 5items
| ||||||||||||||||||
 Citation Citation | Journal: Nat Struct Mol Biol / Year: 2018 Title: Cryo-EM structure of the human neutral amino acid transporter ASCT2. Authors: Alisa A Garaeva / Gert T Oostergetel / Cornelius Gati / Albert Guskov / Cristina Paulino / Dirk J Slotboom /  Abstract: Human ASCT2 belongs to the SLC1 family of secondary transporters and is specific for the transport of small neutral amino acids. ASCT2 is upregulated in cancer cells and serves as the receptor for ...Human ASCT2 belongs to the SLC1 family of secondary transporters and is specific for the transport of small neutral amino acids. ASCT2 is upregulated in cancer cells and serves as the receptor for many retroviruses; hence, it has importance as a potential drug target. Here we used single-particle cryo-EM to determine a structure of the functional and unmodified human ASCT2 at 3.85-Å resolution. ASCT2 forms a homotrimeric complex in which each subunit contains a transport and a scaffold domain. Prominent extracellular extensions on the scaffold domain form the predicted docking site for retroviruses. Relative to structures of other SLC1 members, ASCT2 is in the most extreme inward-oriented state, with the transport domain largely detached from the central scaffold domain on the cytoplasmic side. This domain detachment may be required for substrate binding and release on the intracellular side of the membrane. | ||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 6gct.cif.gz | 223 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb6gct.ent.gz | 178.2 KB | Display | PDB format |
| PDBx/mmJSON format | 6gct.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/gc/6gctftp://data.pdbj.org/pub/pdb/validation_reports/gc/6gct | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  4386MC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |

-Links
 PDBj
PDBj




- Assembly
Assembly
| Deposited unit | 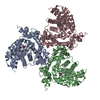
|
|---|---|
| 1 |
|
-Components
| #1: Protein | Mass: 56638.902 Da / Num. of mol.: 3 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: SLC1A5, ASCT2, M7V1, RDR, RDRC / Production host:  Komagataella pastoris (fungus) / References: UniProt: Q15758 Komagataella pastoris (fungus) / References: UniProt: Q15758#2: Chemical | 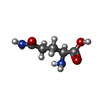  Type: L-peptide linking / Mass: 146.144 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: C5H10N2O3 Type: L-peptide linking / Mass: 146.144 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: C5H10N2O3 |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: human ASCT2 SLC1A5 / Type: COMPLEX / Details: human ASCT2 SLC1A5 / Entity ID: #1 / Source: RECOMBINANT |
|---|---|
| Molecular weight | Value: 0.172 MDa / Experimental value: YES |
| Source (natural) | Organism: Homo sapiens (human) |
| Source (recombinant) | Organism: Komagataella pastoris (fungus) |
| Buffer solution | pH: 7 Details: 20mM Tris-HCl pH 7.4 300mM NaCl 1mM L-glutamine 0.05% DDM 0.005% CHS |
| Specimen | Conc.: 2.5 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Specimen support | Grid material: GOLD / Grid mesh size: 200 divisions/in. / Grid type: Quantifoil, UltrAuFoil, R1.2/1.3 |
| Vitrification | Instrument: FEI VITROBOT MARK II / Cryogen name: ETHANE / Humidity: 100 % / Chamber temperature: 278 K |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Talos Arctica / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TALOS ARCTICA |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 200 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 200 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 49407 X / Calibrated magnification: 49407 X / Nominal defocus max: 2.5 nm / Nominal defocus min: 0.4 nm / Calibrated defocus min: 0.4 nm / Calibrated defocus max: 2.5 nm / Cs: 2.7 mm / C2 aperture diameter: 100 µm / Alignment procedure: COMA FREE |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER / Temperature (max): 90 K / Temperature (min): 70 K |
| Image recording | Average exposure time: 9 sec. / Electron dose: 0.87 e/Å2 / Detector mode: COUNTING / Film or detector model: GATAN K2 SUMMIT (4k x 4k) Details: Freshly purified protein was concentrated using Vivaspin concentrating devices with a molecular weight cutoff of 100kDa to 2-2.5 mg ml-1. 2.8 ul were applied on holey-carbon cryo-EM grids ...Details: Freshly purified protein was concentrated using Vivaspin concentrating devices with a molecular weight cutoff of 100kDa to 2-2.5 mg ml-1. 2.8 ul were applied on holey-carbon cryo-EM grids (Quantifoil Au R1.2-1.3, 200 and 300 mesh), which were prior glow-discharged at 5 mA for 20 s. Grids were blotted for 3-5 s in a Vitrobot (Mark 3, Thermo Fisher) at 20C temperature and 100% humidity, subsequently plunge-frozen in liquid ethane and stored in liquid nitrogen. Cryo-EM data were collected on a 200 keV Talos Arctica microscope (Thermo Fisher) using a post-column energy filter (Gatan) in zero-loss mode, using a 20 eV slit, a 100 um objective aperture, in an automated fashion using EPU software (Thermo Fisher) on a K2 summit detector (Gatan) in counting mode. Cryo-EM images were acquired at a pixel size of 1.012A (calibrated magnification of 49,407x), a defocus range from -0.4 to 2.5 um, an exposure time of 9 sec and a sub-frame exposure time of 150 ms (60 frames), and a total electron dose on the specimen level of about 52 electrons per A2. Best regions on the grid were screened with a self-written script to calculate the ice thickness and data quality was monitored on the fly using the software FOCUS |
| EM imaging optics | Energyfilter name: GIF Quantum LS / Energyfilter upper: 20 eV / Energyfilter lower: 0 eV |
| Image scans | Movie frames/image: 60 / Used frames/image: 1-60 |
- Processing
Processing
| Software | Name: PHENIX / Version: 1.13_2998: / Classification: refinement | ||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM software |
| ||||||||||||||||||||||||||||||||
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C3 (3 fold cyclic) | ||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 3.85 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 184080 Details: A total of 6345 dose-fractionated cryo-EM images were recorded and subjected to motion-correction and dose-weighting of frames by MotionCor2. The CTF parameters were estimated on the movie ...Details: A total of 6345 dose-fractionated cryo-EM images were recorded and subjected to motion-correction and dose-weighting of frames by MotionCor2. The CTF parameters were estimated on the movie frames by ctffind4.1. Bad images showing contamination, a defocus below or above 0.4 and -3um or a bad CTF estimation were discarded, resulting in 4863 images used for further analysis with the software package RELION2.1. About 3000 particles were picked manually to generate 2D references which where improved in several rounds of autopick. A low threshold was used during the final autopick step to ensure that no particles are missed yielding more than a million particles. Particles were extracted with a box size of 240 pixels, and initial classification steps were performed with three-fold binned data. False positives or bad particles were removed in first rounds of 2D classification, resulting in 628,015 particles that were further sorted in several rounds of 3D classification. A map generated from the GltPh structure (PDB ID 3KBC) was used as reference for the first round, and the best output class was used in subsequent jobs in an iterative way. The best 3D class, comprising 184,080 particles from 4859 images, was subjected to auto-refinement, yielding a map with a resolution of 4.26 A before masking and 3.91 A after masking. Particles were further polished in RELION version 2.1 and subjected to another round of 2D and 3D classification resulting in a final dataset of 133,437 particles. The final polished map had a resolution of 4.26 A before masking and 3.85 A after masking. The map was sharpened using an isotropic B-factor of -171 A2, for manual inspection a B-factor of -225 A2 was used. The approach of focused refinement, where the less-resolved detergent micelle was subtracted from the particle images, did not improve resolution. During 3D classification and auto-refinement jobs a C3-symmetry was imposed. To check for conformational heterogeneity of the data, where single protomers within the trimer might adopt a different conformation, 3D classifications with no symmetry imposed were performed at different stages of image processing. We further performed 3D classification on the individual protomers of a single transporter using symmetry expansion and signal subtraction. Both approaches showed no indication of the existence of a different conformation. Local resolution estimates were estimated by RELION. All resolutions were estimated using the 0.143 cut-off criterion with gold-standard Fourier shell correlation (FSC) between two independently refined half maps. During post-processing, the approach of high-resolution noise substitution was used to correct for convolution effects of real-space masking on the FSC curve. Symmetry type: POINT | ||||||||||||||||||||||||||||||||
| Atomic model building | Protocol: AB INITIO MODEL / Space: REAL | ||||||||||||||||||||||||||||||||
| Refine LS restraints |
|
