National Institutes of Health/National Institute Of Allergy and Infectious Diseases (NIH/NIAID)
R01-AI11893
United States
Citation
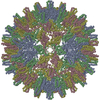
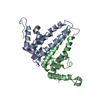
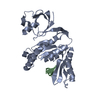
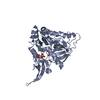
Journal: J Virol / Year: 2019 Title: Structural Differences between the Woodchuck Hepatitis Virus Core Protein in the Dimer and Capsid States Are Consistent with Entropic and Conformational Regulation of Assembly. Authors: Zhongchao Zhao / Joseph Che-Yen Wang / Giovanni Gonzalez-Gutierrez / Balasubramanian Venkatakrishnan / Roi Asor / Daniel Khaykelson / Uri Raviv / Adam Zlotnick / Abstract: Hepadnaviruses are hepatotropic enveloped DNA viruses with an icosahedral capsid. Hepatitis B virus (HBV) causes chronic infection in an estimated 240 million people; woodchuck hepatitis virus (WHV), ...Hepadnaviruses are hepatotropic enveloped DNA viruses with an icosahedral capsid. Hepatitis B virus (HBV) causes chronic infection in an estimated 240 million people; woodchuck hepatitis virus (WHV), an HBV homologue, has been an important model system for drug development. The dimeric capsid protein (Cp) has multiple functions during the viral life cycle and thus has become an important target for a new generation of antivirals. Purified HBV and WHV Cp spontaneously assemble into 120-dimer capsids. Though they have 65% identity, WHV Cp has error-prone assembly with stronger protein-protein association. We have taken advantage of the differences in assemblies to investigate the basis of assembly regulation. We determined the structures of the WHV capsid to 4.5-Å resolution by cryo-electron microscopy (cryo-EM) and of the WHV Cp dimer to 2.9-Å resolution by crystallography and examined the biophysical properties of the dimer. We found, in dimer, that the subdomain that makes protein-protein interactions is partially disordered and rotated 21° from its position in capsid. This subdomain is susceptible to proteolysis, consistent with local disorder. WHV assembly shows similar susceptibility to HBV antiviral molecules, suggesting that HBV assembly follows similar transitions. These data show that there is an entropic cost for assembly that is compensated for by the energetic gain of burying hydrophobic interprotein contacts. We propose a series of stages in assembly that incorporate a disorder-to-order transition and structural shifts. We suggest that a cascade of structural changes may be a common mechanism for regulating high-fidelity capsid assembly in HBV and other viruses. Virus capsids assemble spontaneously with surprisingly high fidelity. This requires strict geometry and a narrow range of association energies for these protein-protein interactions. It was hypothesized that requiring subunits to undergo a conformational change to become assembly active could regulate assembly by creating an energetic barrier and attenuating association. We found that woodchuck hepatitis virus capsid protein undergoes structural transitions between its dimeric and its 120-dimer capsid states. It is likely that the closely related hepatitis B virus capsid protein undergoes similar structural changes, which has implications for drug design. Regulation of assembly by structural transition may be a common mechanism for many viruses.
 Movie
Movie Controller
Controller


 Open data
Open data
 Basic information
Basic information Components
Components Keywords
Keywords Function and homology information
Function and homology information Woodchuck hepatitis B virus
Woodchuck hepatitis B virus X-RAY DIFFRACTION /
X-RAY DIFFRACTION /  Authors
Authors United States, 1items
United States, 1items  Citation
Citation
 Structure visualization
Structure visualization Downloads & links
Downloads & links Other downloads
Other downloads






 PDBj
PDBj

 Assembly
Assembly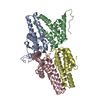
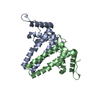
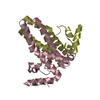

 Sample preparation
Sample preparation Processing
Processing